
-

����ͨ��
����ץס�����Ƽ�
����������
����ͨ��
����ץס�����Ƽ�
����������
RNA�������ص����ص���
�����壺 �� �� С �� ʱ�䣺2021��07��09�� ��Դ������ͨ
�༭�Ƽ���
��������ת¼��ѧԽ��Խ�ܵ���ѧ���ǵ����ӣ���Ϊ�˽����������������֮һ������Ϊֹ����RNA���ѷ�����170���ֻ�ѧ����[1]��
2016������һ�죬Nature Methods���ŷ�����2016��ȼ�������������Ϊ�������ȼ���һ���Ǵ���CRISPR������Ȼ���ѣ������ľ�ǰհ�Ե���ȼ����̵���˵��һ���������������ʣ�Epitranscriptome analysis������ת¼��ѧ���������⡣
Epitranscriptome analysis�����������ϣ���epi����Ϊǰ��ָ�ľ��dz�����֪���ܻ��Ŵ��ԣ��κ����ӵ��������ϵ����Ρ���ʮ��������ѧ���Ǽ�����û��ע�RNA���Σ���Ϊ�����ϸ�����60�����70���RNA�ϵı�Ǿͱ������ˣ����Ǵ��ֻ��ע��tRNA��rRNA���Լ�DNA�ϵı����Ŵ����Ρ�
�����ſ�ѧ���Ƿ����˳���������RNA�����еĻ�ѧ��ǣ���̬���ӻ���ȥ����Щ��ǵġ�Readerд�֡��͡�Eraser��Ƥ���������µ�ȼ�˶�RNA���ε���Ȥ�����磬����������ȥ��һ�������ŵ�ø���밢���ĺ�Ĭ֢��������֮��Ĺ������������������������������������Ҫ�������á�
�ɴ˱���ת¼��ѧԽ��Խ�ܵ���ѧ���ǵ����ӣ���Ϊ�˽����������������֮һ������Ϊֹ����RNA���ѷ�����170���ֻ�ѧ����[1]����Щ���δ����ֲ��ڷDZ���RNA��ncRNA�����ر���rRNA, tRNA��snRNA�ϣ�ΪncRNA�ڷ���������з����������������衣�����˷ܵ��ǣ��о���Ա����m6A��N6-methyladenosine����m1A��N1-methyladenosine����m5C��5-methylcytidine����hm5C��5-hydroxylmethylcytidine����I��inosine���Լ��ף�pseudouridine���Ȼ�ѧ����Ҳ�ֲ����������mRNA�ϣ�Ӱ��mRNA�Ĵ�л�빦�ܡ��ر��ǰ���������mRNA����ø��Writer����ȥ����ø��Eraser��������ʶ�ף�Reader�����·��֣�mRNA��ѧ���εĿ���仯�붯̬�������¼������о���Ա����Ȥ��
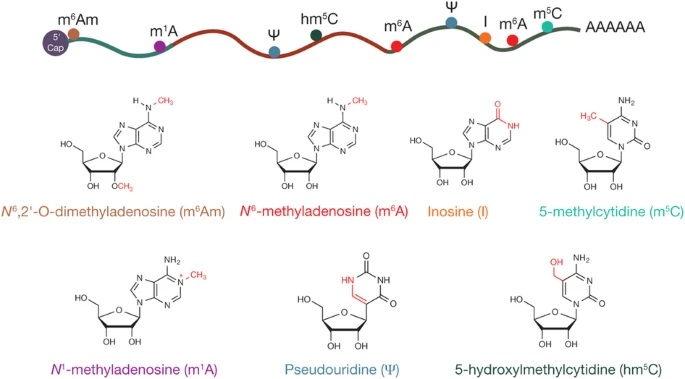
ͼһ����[2]
����ת¼��ѧ�о���Χ
����ת¼��ѧ��epitranscriptomics���ֳơ�RNA�����Ŵ�ѧ������ָת¼�� RNA ���Σ���Щ���θ�ת¼������˹�����صı仯������ת¼�����ΰ���������Ҫ�� RNA �ӹ��¼������� RNA �༭�������ͼ��ӣ�ͼ��)��

ͼ������[4]
���Ƿ���뵰������˵��RNA���Է�Ϊ����RNA��coding RNA�����κͷDZ���RNA��non-coding RNA, ncRNA�������ࡣǰ�߾���ָmRNA������������ܶ����࣬��������֪��tRNA��rRNA������RNA���ε�snoRNA�ȡ�
mRNA�����о����ľ���m6A ���Σ����� 20 ���� 70 �������ѧ���Ǿ��� RNA �з����� m6A ���Σ����Խ��Խ����о�֤��m6A ���ε���Ҫ�ԣ�m6A ���κ� mRNA ���ȶ��ԡ����Ӽӹ��������Լ� microRNA �ļӹ��йأ�m6A ����ϸ�����ˡ����������أ����Դ�ʹ��ϸ�������Ҹ���״̬ת��ϸ���ֻ����о���Ա���֣����������� mRNA �İ�˥�ڣ��������ȡ�����˵��m6A ���μ���Ӱ�� RNA ��л��ÿ�����衣
���������������о������룬�����о�����Ҳ��mRNAתΪ��ע�DZ���RNA�ļ������ڼ���������չ���̵���Ҫ���ã����ַDZ���RNA��m6A�������ڸ�ϸ���ֻ�����ϸ����ֳ�ȹ�������ؼ����á�ͬmRNAһ����lncRNA��Ҳ�����Ŷ��ֻ�ѧ���Σ�m6A��������lncRNA���������Ե���lncRNA�����ṹ��lncRNA��ϵ��ף��Լ�lncRNA��ceRNA���ƣ��л����m6A���Ρ�
����л�״RNA��m6A���Σ�����Ӱ��circRNA��RNA��ϵ���(RBP)֮�������ã��Լ������ԴRNA���Ӷ���������ԴRNA���ֿ������ⱻ��������ϵͳʶ����
Small RNA������microRNA��tRNA��ԴСRNA(tsRNA������tRF&tiRNA)�ȣ���RNA�����Ͼ��ж��ֲ�ͬ�����Σ���Щ����һ�����ܹ�����small RNA�Ļ��ԣ���һ����Ҳ���Ը��������µĹ��ܡ���֪��ЩRNA����ͨ�����ַ��ӻ��������ӹ��ܣ���RNA���ο��Ըı�miRNA�İ����Ի�ı�tsRNA(tRF&tiRNA)��RNA��ϵ�����������������������Եȡ�Small RNA���������DZ���ת¼��ѧ�о�����ǰ�أ�������Ҫ�Ŀ�ѧ������ٴ���ֵ��
��������ΪֹRNA������������Ҫ�ijɾͣ����ζ���ѧSamie R. Jaffrey������Ϊ������һ�����ڻ��� m6Aȫת¼�鷽�������DZ���ת¼��ѧ����������Ҫ���¼��������Ѿ������������������ֺ��������Ρ� ��ȫת¼����ͼ֮ǰ��ͨ��ʹ�����������������������ˮ�� RNA �����εĺ����ᣬ�������εĺ����ᡣ��Щ������ģ�����ɵģ������Ƕ��ڵͷ�ȵ����Σ���ʹ����һ���ߴ��ȵ� mRNA �Ƽ����㻹�ǵ���������Ⱦת�� RNA (tRNA) ������� RNA (rRNA) ���������ε���Դ����ʹ�ú���ȷ�����εĺ������Ƿ����� mRNA����
��ɫ������ά���ѧҽѧԺ��Gideon Rechavi�������ʾ�������ҿ��������������Ҫ�ɾ���ͨ��mRNA ���ν�ʾ��һ���µġ����ӵġ��߶����еġ��ɵ��ڵĻ��������ز㡣��һ�µĵ��ز�������mRNA �Ķ������ԡ��������Ƕ��ݵġ��߶Ƚṹ���ġ���ϸ������֮���ƶ���ͨ��ת¼�Ŵ�ġ���ЩӰ�첿�����ɡ��Ķ������鵼�ģ�����������Խ�ϵ��ף����ļ��������������һ����̱����������ĵ���Ҳͨ����д�֡��͡���Ƥ�������İ�װ��ɾ��֮�������ý��е������ڹ�ȥ��ʮ���У������˼�����Ҫ�Ľ�ѵ�����ȣ�mRNA ���ηdz��ձ飬��ǧ������ת¼�������Ρ���Ȥ���ǣ�һЩ�ľۼ����ض���ת¼λ�ã����磬������Ҫ�������ظ��� Alu ��������m6A ����������ֹ�����Ӹ������ڲ������ӡ� AUG ��ʼ��������Χ�� m1A �أ������ÿ������ͨ����ͬ��ģʽ�������ж������⣬һЩ���Σ��� m6A �� m1A�����˺�С��֮����ֳ��߶ȵı����ԡ�
��һ����Ҫ�ijɾ��Ƿ����ض����Ŀ���ͨ����ͬ����Ϊģʽ��ͨ����ͬ�Ķ�ȡ�������������Ρ�����һ����Ҫ������һЩ mRNA ���εĶ�̬���ԣ����ԶԻ����̼��������ٷ�Ӧ�� m6A �� m1A �Ѿ�֤�������ֶ�̬���ԡ� mRNA ���εĺ��������������쳣���ζ������С�����ڷ����Լ����֢����֢�����Ե��ƻ���Ӱ�죬��һ��ǿ������һ���ڲ����Ҫ�ԡ���
������ѧ��Tsutomu Suzuki���ڱ�ʾ������ǰ���� RNA ���ε������о���Ҫ�����ھ���ķDZ��� RNA������ tRNA��rRNA ��С�� RNA (snRNA)����Ϊ��ЩRNA ��ϸ���к����ḻ��Ȼ���������ʹ�� NGS ������ RNA���ν���ȫת¼������Ѷ������ּ�����Σ����� mRNA �еļ��� (I)��m6A��m5C���� �� m1A �Լ������DZ��� RNA���ⷴ���������ؿ��˱���ת¼��ĸ��
�ڹ�ȥ��ʮ�����У�ͨ�������Ŵ�ѧ����������ѧ����ʶ����������ģʽ����������е�RNA ����ø��ʹ�����ַ��������dzɹ������� 40 ��RNA ���λ���
��ֵ��һ����ǣ��뼲����ص�����������������ڽ���RNA ����øͻ������������������༲������[5]
��Ŀǰ���о��ɹ�����������ת¼��Ӱ����Զ������Ϊ�������Ե���������������������ת¼���ϵ�һ���ֶΡ�����ת¼�顰���롱�����������û���ǿ RNA ���� RNA �����Է�Ӧ�е��ض���ѧ��Ӧ�������Ըı� RNA �ṹ-���ܹ�ϵ���Ӷ���ʱ�պ��ź������Է�ʽ�ṩ����Ļ�����ز㡣
Ȼ��������ת¼��Ĺ����о�����ڱ��ۻ�����Ĺ����о���������Ϊȱ��������ת¼�鷶Χ�ڼ����Щ����ת¼���ǵ��������Ƚ��ļ������о�����ת¼����ڼ�����Ҫ��ս�����ȣ������ RNA ���β���ͨ����ͨ������ֱ�Ӽ�����Ϊ�� RNA �Ļ�ѧ����ͨ������ı����μ���ļ��������ԣ���ת¼ (RT) ���ز�����Щ���Σ���ʹ�����볣�� RNA ��������֡���Σ���Ȼ rRNA��tRNA �� snRNA �ܷḻ�����������͵�RNA������ mRNA �ͳ����DZ��� RNA (lncRNA)����ȿ��ܽϵ͡�������ȱ�����еļ��㹤�����ٽ��Ӳ�������ʶ������λ���������
���˵��ǣ�����������Բ�ͬ����ת¼����о�����ȡ�����ش��չ����Щ�¹��߰����о���Աȷ�� RNA ���ε�λ�ã�����ʾ��Щ����������ת¼���еIJ�ͬ�ֲ�ģʽ������Щ�������������˹��ߣ����������༭���ߣ����ʹ��ʱ��RNA ����ø�İб����ȷ�������⣬��Щ��������ʾ�˲�ͬ���������²�ͬ����ת¼���ǵĶ�̬���ʡ�ͬʱ��Щ����ʹ�����ܹ�����ѡ����ʶ���ض�����ת¼���Dz�ȷ���书�ܵġ��Ķ��������ס���ˣ��µ��о����߲������ԶԱ���ת¼�����ȫ����������һ��ǹ��ܱ���ת¼���о��ı�����Դ [2]��
�� ����������
�о� RNA ���εķ���������Һ��ɫ��(LC-MS) �����ڸ�ͨ������ķ�����оƬ�����ȷ�����
�� mRNA&lncRNA����ת¼��оƬ
RNA���ε�DZ�ڹ��ܲ���ȡ�����������ε��Ǻ��ֻ���ת¼����ͬʱҲȡ���ڱ����β����ڸ�ת¼������ռ�İٷֱȡ�Ȼ����Ŀǰ��ת¼��ˮƽ�ĵ�RNA���μ�ⷽ��������Ѱ��ת¼���ϵ�����λ�㣬���ܹ������ؼ�ⱻ����ת¼���İٷֱȡ���һ�ඨ����Ϣ��ȱ��������Խ��Խ����й����ߵĹ�ע��
mRNA���ε�DZ��Ӱ���ȡ���������ЧӦ��Ҳȡ���ڱ�����ת¼���İٷֱȡ����磬һ�ֿ��Լ���mRNA��������Σ����ֻ��1%��ת¼�������Σ���Ȼ��̫���ܲ����κ����﹦�ܣ�Ȼ����һ�����ο��Դ�ʹmRNA������µĵ�������ʱ����ʹ����ˮƽ�ܵͣ�Ҳ���ܲ�����Ҫ�����﹦�ܡ���ǰ��m6A�ͦ���ⷽ������������ȱ�����γ̶ȵĶ�����Ϣ��m6A�ĵ������ÿ���ͨ��pulldown m6A�ķ���������ض������ڲ�ͬ״̬�µ���Ը����̶Ƚ����ƶϣ������ܴ���Щ�����л�ñ�����mRNA�ľ��������µĿɶ���m6A�ͦ��ĸ�ͨ����ⷽ����������Ĵٽ�������ķ�չ[7]��
��һ����Ҫ�������Dz���RNA���εĻ�ѧ�����Ķ�̬�仯��Ŀǰ������ת¼��ѧ�о����ص�������Щλ�㱻���Σ�������RNA��ÿ��������λ���ռ�ȡ���ͨ������mRNA�Ͳ���RNA��m6A����λ����ʾ���κ�m6A����λ���ռ�ȶ�����ﵽ100%�����εĻ�ѧ�����仯������һ��RNA����ѧ���εĶ�̬�仯���������ο���Ӱ��mRNA�Ľṹ���ͣ���RBPs����ļ���κ�����λ������Σ��ᵼ��ͬһmRNAȺ��������ڽṹ���ǽ��reader�IJ�ͬ����Ϊ����mRNA��Ⱥ����ˣ��ı����εĻ�ѧ��������������ͬһ��RNAת¼����ʹ��ͬ���ܵ�һ�ֻ��ơ�Ŀǰ������Լ�����λ�ѧ�������ĸ�ͨ���������Բ�������ת¼��ѧ����һ���������[8]��
Arraystar mRNA&lncRNA���۹�ת¼��оƬ�����˫ӫ��ͨ��оƬ������RNA�������߹�������������ת¼��ˮƽ��RNA���ν��ж�����⡣�����ı���ת¼��ͼ��ΪRNA���ε����о��ṩ��Ҫ��Ϣ��
���⣬����Arraystar small RNA����оƬ�������ṩ����ؼ��������ڵ���оƬ�ɶ���miRNA��pre-miRNA��tRNA������small RNA��tsRNA������tRF��tiRNA���ļ�����Ρ��ɼ������ΰ�����8-���������ʣ�o8G����7-�����գ�m7G����N6-�����գ�m6A���������գ�������5-�����գ�m5C����
��ȡ��������Arraystar ����ת¼��оƬ
��Һ��ɫ��-����
LC-MS ��һ��ǿ��ļ��������Գ�ɫ�������Ⱥ������Լ�����ι���RNA���ա�Ȼ�����ڲ��� mRNA ���εı����£��������ؼ����ơ�
���ȣ�LC-MS ����λ���ض������������ޡ��� mRNA �ı����£�����Ϊֹ��MS ����������ȫ�����ĺ��գ�����ܹ�������Ʒ�����εġ����塱ˮƽ�����ų��˽������η��䵽����λ��Ŀ����ԡ���Ӧ���ڲ��������� RNA �Ѻ�����ʱ��LC-MS ԭ���ϻ������ṩλ���ض���Ϣ��Ȼ�������ַ���ͨ����Ҫ��ʮ�������ɿ˵����ٲ��ִ����ķ��ӣ������ mRNA ��˵�Dz���ʵ�ģ���Ϊ�����и߶������Ժ͵ͷ�� �����ԡ�
��Σ��� tRNA �� rRNA ��ȣ�mRNA �����ε����ˮƽԽ�ͣ���˽��� LC-MS ������Խ��Խ���ѡ���ʹ���Ը߶ȱ���� tRNA �� rRNA �ĵ�ˮƽ��Ⱦ����Զ����ȫ���⣬�������ǿ�����һ���̶��ϻ��� tRNA �����Ի� rRNA �����Ժ��ս��й��ƣ���Ҳ���ܵ��¶�mRNA����ˮƽ�ĸ߹���m6A �� MS �㷺���еĵ�һ��mRNA ���η��������ⷽ��ʩ�ӵ����ƽ��٣���Ϊ���� mRNA �и߶ȷḻ������ tRNA �в����ڣ����ҽ������� rRNA �е�����λ�㣨��ˣ���Ⱦ���ܵ��µ�ʵ�� m6A ˮƽ����
���֮�£���������� m6A ����ת¼���� mRNA �ж��dz��������� tRNA �� rRNA1 �и�Ϊ�ձ飬������������ LC-MS �˽����ȺͶ�̬����������һЩ����£�ͨ�� LC-MS ���Ƶ����η�ȣ����ڸ����α���Ϊ�� mRNA ���Ը�ˮƽ���ڣ��ͻ��ڻ����鷽����ͨ������� mRNA �еĴ������Σ�����ˣ���Щ����ĸ�Դ�������� mRNA �����е� tRNA �� rRNA ��Ⱦ���һ�ֿ�������ijЩ������ RNA ���˵Ľ��������ܵ��º��������������������������ɢ��������ת¼���еķ�ø�����ο���ͨ�� LC-MS ����������ͨ�������鷽����������Ҫ���ض�λ������źš�
�ۻ��ڲ���ķ���
�ڹ�ȥʮ���У����ڸ�ͨ������������������Ѿ���Ϊ�˸���������Ĺؼ�����������Щ�������Ը�֪ mRNA �����εĴ��ڼ�����ת¼���еľ�ȷλ�á�
�� LC-MS ��ȣ�ԭ���ϵ����������̾Ϳ��Ի�֪�������εķ�ȣ���ʹ�û����鷽��ʶ��������ҪΪÿ�����ο���ר���Ҷ��صĹ������̡����ַ�����Ҫ�˷��ĸ�����ս�ǣ�����������ڱ��������ǡ����ɼ��ġ���Ҳ����˵�������ڽ� RNA ��ת¼Ϊ cDNA �������κκۼ������ǻ��� Illumina �ı�������Ⱦ����跽������ˣ���ѧ�����Ѿ�������һ���������ֲ�ͬ�Ŀɼ��ġ����� m6A����Ҫ��ⷽ�������ڿ� m6A ���壬��Щ��������ѡ�������߳������� m6A �Ķ� RNA Ƭ�Ρ�
�������߳�����һ��ͨ�÷�����ԭ���Ͽ������ڼ���κ����Σ���ʵ���ϣ����ڿ���ķ����ڻ��Ʒ� m6A ����ת¼���ͼ�����Ч���൱���ޡ������о���Ա����ʹ���������� m1A �� ac4C36 ��ͼ�ף����ڱ���Ϊ�ᵼ�´���������λ�㣬����������ڿ��彻�淴Ӧ��ɵġ��� m6A ����Ҳ�������ֽ��淴Ӧ�ԣ�Ȼ�������� m6A �ķ����Խϸߣ��������Ȼ�ɿء����֮�£��Խϵ��������ķ�ȴ��ڵ����ε��·��������������Խ���¼��ı��ʼ������ӡ�
��ˣ����ھ�������� m6A ����ת¼�飬�Ѿ��������������μ���Ķ��ػ�ѧ����ʹ��������ת¼��ɼ��ķ���������ͨ����ͬ�IJ���ʵ�ֵģ����磬ͨ���ı����μ�����ݵ����������Ի�ѧ���ʻ�ͨ���������л��ض����ӵ����μ���ϵ�����ת¼�����й���ضϣ����±�������Щʵ�鷽��ͨ���ᵼ����λ��Ĵ����ȱʧ�����ߵ��¶�ȡ�ġ��ѻ�������Щ��ȡѡ���Ե����ض�λ�ÿ�ʼ�������Ȼ����Դ����ƶϳ��ĵĴ��ڡ����ѧ������ѧ��������Ҫ�����������ɫ�ġ�ͨ��Ϊ��������ķֱ��ʣ��Լ������ܹ��ṩ����ˮƽ����Զ�������ʱ�����Ǿ��Զ���[1]��
���翵�����ィ����һ�ָ������ľ��Զ�����ʵʱӫ�ⶨ��PCR�������������������ڵ�������ֱ���ˮƽ��ȷ���RNA��m6A����λ��, ����δ֪ģ����ж����������÷����ɳɹ���Ӧ����ʵ��������Ʒ��m6A���εľ�ȷ��⣬��ʹ�ǵͷ��RNA��������
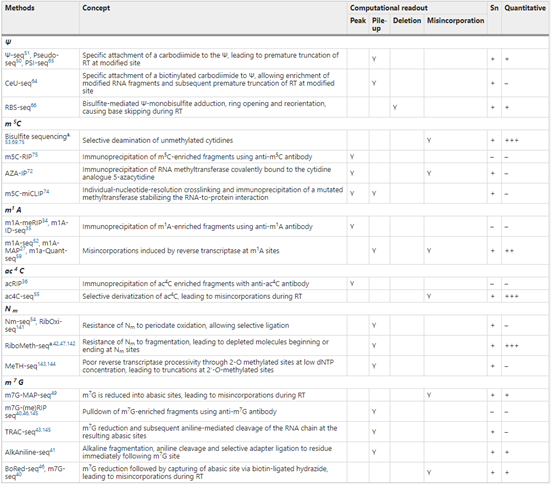
������Ҳ��һЩ�µIJ�����½�����֣�����2017 �꣬������ѧ���������������ɫ��κ�����о������о��Ŷӷֱ���������˵�����ֱ���ˮƽ��� m1A ���·�������m1A-MAP ��m1A-Seq��2019 �֥꣬�Ӹ��ѧ Bryan C�� Dickinson �Ŷ���δ��ŶӺ������о������˿ɿ���ѡ����ת¼ø�Ľ���ƽ̨�������ø�ƽ̨�о� mRNA �ϵ� m1A ���Σ�ʵ���˵����ˮƽ m1A �ļ�⡣
���⣬ֱ�ӵ�RNA���ײ���Ҳ���ܹ�ע��RNA�����Ŵ�ѧ������������ζ���ѧ��Christopher Mason��ʾ������ȥ������ͨ�������ÿ����ѧ�������ƶ�RNA������״̬��ֱ����������Dzſ�ʼ����ֱ�ӵ�RNA�����������ײ����ǣ������ܹ�ֱ�Ӳ���RNA��������Ҫ��ת¼�����ǵ�һ��ֱ�ӲⶨRNA���Σ��������Ӵ�����Щ���Σ��ִ�����Щ�칹�塣��
���Կ���˵�����ײ���������������RNA���е�m6A,m5C���Σ�һ���Ƕ�ȡ����������ʱ�������μ�����ӳ٣�����������ʹ֮�Կɱ�����о����칹��������ƣ�����ƴ�ӣ�Ҳ������ʧ�������ӣ����Ǿ��Ƿ��ñȽϹ�
�������һƪ�ȵ����¡�The Architecture of SARS-CoV-2 Transcriptome SARS-CoV-2���У��о���Ա����MinION���ײ����ǽ�����DRS�������879,679��reads��1.9 Gb�����������֤���˲���ת¼��ռ��������λ����������DRS����RNA�ĵ����Ӽ�⣬�ṩ�˶��صĻ�������鵥��RNA���ӵĶ��ת¼���������������¹ڲ���RNA�����ص�[9]��
��Ŀǰ��֪��RNA����ת¼���Σ�
m6A
m6A���������mRNA�Ϻ�����ḻ�Ļ�ѧ���Σ��ɼ�ת��ø���������METTL3��METTL14��WTAP��KIAA1429��RBM15��RBM15B�����������ɱ�ȥ����øALKBH5��FTOȥ����Ŀǰ�ѷ����˶���������ʶ��m6Aλ��ĵ��������YTH���嵰�ף�YTHDF1-3��YTHDC1����ת¼��ʼ������eIF3�����Ǻ˵��ף�HNRNPA2B1��HNRNPC���Լ�RNA��ϵ���SRSF2��m6A��Ҫ�ֲ�����ֹ�����Ӹ�����3��UTR����Ӱ��RNA��ԡ��ı�RNA�����ṹ������ֱ��ʶ�𣬽�������mRNA�ij��졢�ɱ���ӡ��ȶ��Ժͷ�����̡���A-U�����ȣ�m6A-U��Խϲ��ȶ�������RNA�ڲ�˫������������ṹת�䡣m6A����˫���뵥���Ĺ�������ѵ�����ǿRNAת�������ȶ��ԡ�ȥ���������ʹmRNA�ָ�ԭ���������ֹ���ת����ܵ���mRNA�벻ͬ��������øı䣬�Ӷ�������ͬ������ѧЧӦ��m6A��ֱ�ӱ��ض�������ˮ�ṹ����ʶ�𡣱��磬YTH���嵰���������Ե�ʶ��m6A���ر���GGm6ACU�������С����Ա֮һYTHDC1ʶ���m6A�����ذ���mRNA�Ŀɱ���ӡ�����һ��ԱYRHDF2��m6A��Ϻ���ļCCR4-NOT������ٽ�����RNA�Ľ��⡣��UV��������ݿ˷�Ӧ�У�ת¼��ʼ������eIF3��5��UTR����m6A��ϣ��ٽ�ñ�������ķ�����̡���������m6A�ɱ�SRSF2ʶ�𣬲���֬�����ɵĵ��ء��ڹ�Ӭ�У�YTHDC1ͬԴ����ʶ���Ա�����mRNA�ϵ�m6Aλ�㣬������ɱ���ӣ��Ӷ����ƹ�Ӭ���Ա���ǰ������m6A�鵼��mRNA�ȶ��Ե���Ҳ�Ը�ϸ���ֻ����������ʱ�ӿ���ʮ����Ҫ�����⣬m6AҲ��ͨ��Ӱ��mRNA��tRNA�������ӵ���������뱣��ȣ��Ӷ�Ӱ�췭�����졣
�о��������棬Arraystar m6A������ֱ���оƬ����RNAøMazF��m6A���е����ԣ����ڵ�����ֱ���ˮƽ��λm6Aλ�㣬��������ж�������оƬ���������ִ漼�����������ƣ����о�m6A��̬�仯������ѧ�����Լ���������Ե�ǿ�ߡ�
m1A
m1A���½����ֵĿ���ı���ת¼���Σ��ܱ�RNA��øALKBH3ȥ����Ŀǰ��δ������ȷ��m1A����ø������ʶ�ס���m6A��ͬ��m1A�ı����Ƚϵͣ���Ҫ�ֲ���mRNA��5��UTR�������ܲ�����ڷ�����ʼ���̡�m1A����ȫ��ֹWatson-Crick��ԣ�����RNA˫�����������ٽ�RNA-���ľ�������û�RNA�ɱ�����ṹ���γɡ�m1A������ѧ��������δ֪�����о����֣������ݿ˻�Ӫ���ѷ���ѹ�������£�ϸ���ڵ�m1A����ˮƽ������������ͨ���ٽ�ñ�����Է��룬����ϸ����Ӧ����Ӧ��
m5C��hm5C
m5C�㷺�ֲ���tRNA��rRNA�ϣ������ȶ�tRNA�����ṹ��Ӱ�췴�����ӻ�����ά��rRNA���뱣��ȹ��ܡ��½���RNA���������֣�mRNA�ı�������DZ������ϴ���8000���m5Cλ�㣬�����൱һ����λ�㼯����5��UTR��3��UTR����m5C���ɼ�ת��øNSUN2��TRDMT1���γɣ���˫����øTET�����γ�hm5C��hm5C���ܾ�����һ�������γ�f5C��������ذ���ऺ��գ�C����m5C��Ӱ������ԣ���������ǿ����ѵ��Լ�RNA�뵰����ˮ���á�m5C���ж�������ѧ���ܡ�p16 mRNA�ڱ�NSUN2ø����m5C���κ��併�ⱻ���ƣ��ȶ�����ǿ����ϸ�������У�NSUN2�ı����ܵ����ܵ��ء�NSUN2����CDK1 3��UTR������m5C���Σ��ٽ��䷭�룻ͬʱ��CDKN1B��5��UTR������m5C���Σ�����CDKN1B�ķ��롣���߹�ͬ���ã���ǿϸ������ֳ������m5C����˥����ػ���ķ�������йأ�������NSUN2�����ӻ�������˥�ϵķ�������Ϊm5C���������hm5CҲ����ǿ����Ч�ʡ�hm5C�ڹ�Ӭ�����б������ܸߣ����ܲ����Ӭ���Բ�������
Pseudouridine (��)
������ऺ��զף�������Ϊ����������ᣬ������ऺ��գ�U���칹���γɡ����˵�ϸ����С����֯��mRNA�У���/U�ı���ԼΪ0.2-0.6%��������������व��칹����Ӧ��PUSø����������H/ACA���Ǻ˵���һ�𣬴���ɡ���������ܼ���RNA����Ŀɱ��ԣ���ǿ�������ȶ����Լ��뵰��֮��ĵļ�������á�������ि��ܵ���mRNA�ȶ��Ժͻ����������ĸ�����ݿ˷�Ӧ�����������Ŀǰ�в���ȷ��
Inosine (I)
�������Σ�����ΪA-to-I�༭���Ǹߵ���������������һ��RNA�༭��ʽ�����������Ѱ�øADAR��ɡ�A-to-I�༭��Ҫ�����ڷDZ��������ں�������AluԪ���ڡ�A-to-I�༭��ȫ�ı��˼��������ԣ�AU���ת��ΪIC��ԣ��Ӷ��ı�������İ����ᡣ���磬A-to-I�༭�����ԹȰ��������еĹȰ������ر���Ϊ�����ᣬ�����˸�����ͨ�Եĸı䡣���⣬A-to-I�༭�����иı�ɱ���ӡ�����miRNA�IJ����빦�ܡ��Լ�������������߷�Ӧ�����á�
ac4C
2018�꣬����NIH��Shalini Oberdoerffer������Cell�����о����ģ�Acetylation of Cytidine in mRNA Promotes Translation Efficiency���״ν�ʾmRNA�ϴ��ڴ���ac4C���Σ�����ac4CӰ��mRNA���ȶ����뷭��Ч�ʡ�
N4-acetylcytidine (ac4C)��N4λ��������ण������ԭ�������б��صĻ�ѧ���Σ������о���Ϊac4C��Ҫ����tRNA��18S rRNA�ϡ��������о���ʾ��mRNA��Ҳ���ڴ�����ac4C����������������mRNAЯ����m7Gñ�����Ρ�NAT10��Ŀǰ������Ψһͬʱ����������ø�ṹ���RNA��Ͻṹ��ĵ��ף���˱���Ϊ��RNA ac4C����ø��
�ɴˣ���������Ϊһ���µ� mRNA ���Σ�������ת¼����չ����֪���ļ������칹��֮�⡣��Щ��������չ�� mRNA ���ε���֪�⣬��ȷ���˶��ֻ�ѧ���εĴ��ڣ�mRNA ���ε���ģʽ��δ���ܿ��ܻ�������֡���һ���˽���Щ���εĹ��ܡ������������ض�ת¼���ϵĴ��ڵ�;���ͻ��ƣ��Լ������ڸ��������ͼ��������µĹ��ܣ��������ڸ�ȫ����˽����ת¼���ڻ�������е�����[6]��
TGM����
5���˳�������TMG���������类���ֵ�����֮һ�������౻RNA�ۺ�øIIת¼��RNA�У������ǷDZ���RNA��������������Ҫ��ɳɷ�snRNA��5���˵�m7G��ñ�ӻᱻ���������γɾ�����������N2, N2, 7-�������գ�TMG��ñ�ӣ��㷺��������������С��������պϳ�ø��Tgs1�����������������Ѿ���֤ʵ�Ǻϳ�TMGñ�ӵ�Ψһ��ø���ڽ����ϴӽ�ĸ������ܱ��ء���������ʵ��ľ����ԣ�TMG���ζԼ�����RNA�ĵ��ػ��ƻ����ںܶ����ĵط��������ڴ������ϸ��ģʽ�����ж���ֱ�Ӵ����ڶ�TMG���ν����о�����ˣ�TMG�����Ѿ�����������ʮ���꣬�������ڶ�ϸ�������еĹ��ܲ�����ȷ��
������ͨ�����ƣ�
�ο����ף�
1. The epitranscriptome beyond m6A,Nature Reviews Genetics volume 22, pages119�C131 (2021)
2. Epitranscriptome sequencing technologies: decoding RNA modifications,Nature Methods volume 14, pages23�C31 (2017)
3. Detecting RNA modifications in the epitranscriptome: predict and validate,Nature Reviews Genetics volume 18, pages275�C291 (2017)
4. RNA editing-dependent epitranscriptome diversity in cancer stem cells,Nature Reviews Cancer volume 17, pages381�C392 (2017)
5. RNA modifications: what have we learned and where are we headed? Nature Reviews Genetics volume 17, pages365�C372 (2016)
6. mRNA acetylation: a new addition to the epitranscriptome��P. Cody He & Chuan He Cell Research volume 29, pages91�C92 (2019)
7. Wendy V. Gilbert, Tristan A. Bell, Cassandra Schaening. Science (2016)
8. Cole J.T. Lewis, Tao Pan, Auinash Kalsotra. Nat Rev Mol Cell Biol (2017)
9. The Architecture of SARS-CoV-2 Transcriptome,Cell,April 23��2020
����ͨ�Ź��ں�
���ѣ�����ת¼��|RNA����|�DZ���RNA����|
֪����ҵ��Ƹ

