
-

ЙъОпНЁ№ЩОў
ЕгДгЧҘЧЎЙъГьҝЖјј
Мш¶ҜөДВцІ«
ЙъОпНЁ№ЩОў
ЕгДгЧҘЧЎЙъГьҝЖјј
Мш¶ҜөДВцІ«
Т»ЦЦРВөД»щТтұнҙпөчҝШ·ҪКҪЈәРЎ·ЦЧУRNA[ҙҙРВјјЗЙ]
ЎҫЧЦМеЈә ҙу ЦР РЎ Ўҝ КұјдЈә2003Дк12ФВ19ИХ АҙФҙЈә
ұајӯНЖјцЈә
ЎЎЎЎ
RNAТ»¶Иұ»ИПОӘҪцҪцКЗDNAәНө°°ЧЦКЦ®јдөДЎ°№э¶ЙЎұЈ¬ө«ФҪАҙФҪ¶аөДЦӨҫЭЗеіюөДұнГчЈ¬RNAФЪЙъГьөДҪшіМЦР°зСЭөДҪЗЙ«Ф¶ұИОТГЗФзЗ°ЙиПлөДёьОӘЦШТӘЎЈRNAёЙИЕЈЁRNA interferenceЈ©өД·ўПЦК№өГИЛГЗ¶ФRNAөчҝШ»щТтұнҙпөД№ҰДЬУРБЛИ«РВөДИПК¶Ј¬ёьТтОӘҝЙТФјт»Ҝ/Мжҙъ»щТтЗГіэ¶шіЙОӘСРҫҝ»щТт№ҰДЬөДУРБҰ№ӨҫЯЈ¬ТтҙЛёсНвТэИЛЧўТвЈ¬ФЪ2002Дк¶ИScienceЖАСЎөД10ҙуҝЖС§іЙҫНЦРRNAiГыБР°сКЧЎЈЛжЧЕ¶ФРЎ·ЦЧУRNAСРҫҝөДІ»¶ПЙоИлЈ¬СРҫҝИЛФұҝӘКјИПК¶өҪЈәРЎ·ЦЧУRNAөДКАҪзТ»өг¶јІ»РЎЎЈУРИЛНЖІвЈәРЎ·ЦЧУRNAҝЙДЬҙъұн·ўПЦБЛТ»ёцРВІгҙОЙПөД»щТтұнҙпөчҝШ·ҪКҪЎЈЙъОпНЁФзФЪИҘДкөЧөҪҪсДкіхТСҫӯУРәЬ¶аОДХВҪйЙЬУР№ШsiRNAsТФј°ЖдФЪRNAёЙИЕЦРөДЧчУГЈЁІОјыЧЫКцЈәRNAi»Ш№ЛЈ©ТФј°СРҫҝУҰУГ·Ҫ·ЁЈ¬ФЪХвАпЙъОпНЁјМРшОӘОТГЗөДУГ»§ҪйЙЬБнТ»ЦЦФҪАҙФҪТэИЛЧўДҝөДРЎ·ЦЧУRNAЎӘЎӘmicroRNAЎЈ
КІГҙКЗmiRNA
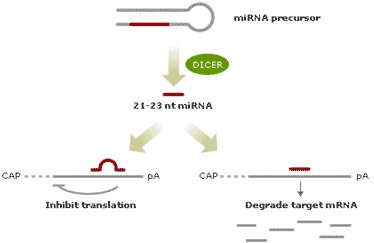
MicroRNAs (miRNAs)КЗТ»ЦЦҙуРЎФј21ЎӘ23ёцјо»щөДөҘБҙРЎ·ЦЧУRNAЈ¬КЗУЙҫЯУР·ўјРҪб№№өДФј70-90ёцјо»щҙуРЎөДөҘБҙRNAЗ°Меҫӯ№эDicerГёјУ№ӨәуЙъіЙЈ¬І»Н¬УЪsiRNAЈЁЛ«БҙЈ©ө«КЗәНsiRNAГЬЗРПа№ШЎЈҫЭНЖІвЈ¬ХвР©·ЗұаВлРЎ·ЦЧУRNAЈЁmiRNAsЈ©ІОУлөчҝШ»щТтұнҙпЈ¬ө«Жд»ъЦЖЗшұрУЪsiRNAҪйөјөДmRNAҪөҪвЎЈөЪТ»ёцұ»И·ИПөДmiRNAКЗФЪПЯіжЦРКЧҙО·ўПЦөДlin-4 әНlet-7Ј¬Лжәу¶аёцСРҫҝРЎЧйФЪ°ьАЁИЛАаЎў№ыУ¬ЎўЦІОпөИ¶аЦЦЙъОпОпЦЦЦРјшұріцКэ°ЩёцmiRNAsЎЈ
miRNA өДМШХч
ТСҫӯұ»јш¶ЁөДmiRNAsҫЭНЖІвҙу¶јКЗУЙҫЯУР·ўјРҪб№№ЎўФј70ёцјо»щҙуРЎРОіЙ·ўјРҪб№№өДөҘБҙRNAЗ°Меҫӯ№эDicerГёјУ№ӨәуЙъіЙөДЈ¬УР5ЎҜ¶ЛБЧЛб»щәН3ЎҜфЗ»щЈ¬ҙуРЎФј21ЎӘ25ntөДРЎ·ЦЧУRNAЖ¬¶ПЈ¬¶ЁО»УЪRNAЗ°МеөД3ЎҜ¶Л»тХЯ5ЎҜ¶ЛЎЈ
ЧоҪь3ёцСРҫҝРЎЧй·ЦұрҙУПЯіжЎў№ыУ¬әНHelaПё°ыЦРјш¶ЁөД100ёцРВmiRNAsЦРЈ¬УР15%ҝзФҪПЯіжЎў№ыУ¬әНІёИй¶ҜОп»щТтЧйҫЯУРёЯ¶ИөДұЈКШРФЈЁЦ»УРУР1ЎӘ2ёцјо»щөДЗшұрЈ©Ј¬Lau әНBartel КөСйКТөДН¬КВёьјУИПОӘЈәЛщУРөДmiRNAsҝЙДЬФЪЖдЛыОпЦЦЦРҫЯУРЦұПтН¬ФҙОпЈЁOrthologЈ¬ЦёДЗР©ЖрФҙУЪН¬Т»ЧжПИЈ¬ФЪІ»Н¬ЙъОпМеЦРРРК№Н¬Т»№ҰДЬөД»щТтИәҫНҝЙұИЧчОӘТ»ёцГЕАаЈ¬ХвР©АаЛЖөД»щТтұ»іЖОӘЎ°ЦұПтН¬ФҙОпЎұЈ©ЎЈ
Bantam ЧоФзұ»ИПОӘКЗ№ыУ¬ЦРІОУлПё°ыФцЦіөДТ»ёц»щТтО»өгЎЈТСЦӘјёёц°ьә¬ФцЗҝЧУөДЧӘЧщЧУІеИлҝзФҪХвёцО»өгөДТ»¶О12.3kbЗшУт»бөјЦВ№ыУ¬өДСЫәНібЦШёҙЙъіӨЈ¬¶шУЙЧӘЧщЧУҪйөјөДТ»¶ОҝзФҪёГО»өгөД23kbЖ¬¶ПИұК§ФтөјЦВН»ұд№ыУ¬ёцМеРЎУЪТ°ЙъРН№ыУ¬ЎЈCohenәНН¬КВУГТ»¶О3.85kbөДЖ¬¶ПөјИл21kbЖ¬¶ПИұК§өД№ыУ¬ЦРК№Жд»ЦёҙФӯАҙөДҙуРЎЎЈө«КЗЖж№ЦөДКЗұнҙпХвёц3.85kbЖ¬¶ПЦРөДESTИҙГ»УРН¬СщөДР§№ыЎЈCohenҪ«ХвёцЖ¬¶ПәНЕұОГAnopheles gambiaeөДН¬ФҙРтБРҪшРРұИҪПЈ¬·ўПЦТ»¶О90bpөДёЯ¶ИұЈКШЗшЈ¬ҫӯ№эRNA folding program (mfold)·ўПЦХвёцұЈКШРтБРҝЙТФРОіЙ·ўјРҪб№№Ј¬К№өГХвёцЗш¶ОәЬПуКЗТ»ёцmiRNAөДЗ°МеЎЈХвёцҪб№ыҫӯ№эNorthern blotЦӨКөН»ұд№ыУ¬өДУЧМеИұЙЩТ»ёц21bpөДbantam miRNA Ј¬УГХвёц90bpөДmRNAЗ°Меҫӯ№эТ»ПөБРөДЎ°№ҰДЬИұК§ЎұЎӘЎ°№ҰДЬ»ЦёҙЎұКөСйЈ¬ЦӨКө bantam miRNAФЪПё°ыФцЦіЦРөДЧчУГЎЈСРҫҝИЛФұУГјЖЛг»ъіМРтјмЛчФЪhid mRNAөД3ЎҜ·ЗұаВлЗшХТөҪБЛbantamөД3ёцЗұФЪөДҪбәПО»өгЈЁ hidКЗ№ыУ¬ЦРТ»ёцУХөјөтНцөД»щТтЈ©Ј¬ІўЦӨКө bantam miRNAТЦЦЖhid өД·ӯТл¶ш·ЗЧӘВјЎЈ
miRNAsөДұнҙп·ҪКҪёчІ»ПаН¬ЎЈІҝ·ЦПЯіжәН№ыУ¬өДmiRNAФЪёчёц·ўУэҪЧ¶ОөДИ«ІҝПё°ыЦР¶јУРұнҙпЈ¬¶шЖдЛыөДmiRNAФтТАҫЭДіЦЦёьОӘСПҪчөДО»ПаәНКұПаөДұнҙпДЈКҪЈЁa more restricted spatial and temporal expression patternЈ©ЎӘЎӘФЪІ»Н¬ЧйЦҜЎўІ»Н¬·ўУэҪЧ¶ОЦРmiRNAөДЛ®ЖҪУРПФЦшІоТмЎЈ
miRNAөД№ҰДЬ
¶ФmicroRNAs (miRNAs)өДСРҫҝХэФЪІ»¶ПФцјУЈ¬ФӯТтКЗҝЖС§јТҝӘКјИПК¶өҪХвР©ЖХұйҙжФЪөДРЎ·ЦЧУФЪХжәЛ»щТтұнҙпөчҝШЦРУРЧЕ№г·әөДЧчУГЎЈФЪПЯіжЈ¬№ыУ¬Ј¬РЎКуәНИЛөИОпЦЦЦРТСҫӯ·ўПЦөДКэ°ЩёцmiRNAsЦРөД¶аКэҫЯУРәНЖдЛыІОУлөчҝШ»щТтұнҙпөД·ЦЧУТ»СщөДМШХчЎӘЎӘФЪІ»Н¬ЧйЦҜЎўІ»Н¬·ўУэҪЧ¶ОЦРmiRNAөДЛ®ЖҪУРПФЦшІоТмЈ¬ХвЦЦmiRNAsұнҙпДЈКҪҫЯУР·Ц»ҜөДО»ПаРФәНКұРтРФЈЁ differential spatial and temporal expression patternsЈ©Ј¬МбКҫmiRNAsУРҝЙДЬЧчОӘІОУлөчҝШ»щТтұнҙпөД·ЦЧУЈ¬Тт¶шҫЯУРЦШТӘТвТеЎЈ
өЪТ»ёцұ»И·ИПөДmiRNAЎӘЎӘФЪПЯіжЦРКЧҙО·ўПЦөДlin-4 әНlet-7Ј¬ҝЙТФНЁ№эІҝ·Ц»ҘІ№ҪбәПөҪДҝөДmRNA°РөД3ЎҜ·ЗұаВлЗш(3ЎҜUTRs)Ј¬ТФТ»ЦЦОҙЦӘ·ҪКҪУХ·ўө°°ЧЦК·ӯТлТЦЦЖЈ¬Ҫш¶шТЦЦЖө°°ЧЦКәПіЙЈ¬НЁ№эөчҝШТ»Чй№ШјьmRNAsөД·ӯТлҙУ¶шөчҝШПЯіж·ўУэҪшіМ(reviewed in Pasquinelli 2002)ЎЈ
bantam miRNAКЗөЪТ»ёцұ»·ўПЦУРФӯ°©»щТтЧчУГөДmiRNAЎЈіэБЛlin-4Ўўlet-7Ј¬ТСЦӘ»№УРТ»Р©miRNAsҝЙДЬІОУлФЪПё°ы·Ц»ҜәНЧйЦҜ·ўУэ№эіМЦРЖрЦШТӘЧчУГөД»щТтөДЧӘВјәуөчҝШЈ¬АэИзmir-14Ўўmir-23 өИЎЈ
ФЪЦІОпmiRNAsөДСРҫҝЦРУРБҪМхПЯЛчМбКҫmiRNAsҝЙДЬІОУлЦІОпөД·ўУэ№эіМЎЈТ»КЗФЪcarpel factory (car) Н»ұдЦкЦР3ёцmiRNAsөДұнҙпЛ®ЖҪПФЦшПВҪөЎЈCARPEL FACTORY КЗТ»ёцАаЛЖDicerөДГёЈ¬ІОУлЦІОпөД·ўУэЈ¬ЖдИұК§Н»ұдЦкұнПЦОӘЕЯМҘәНТ¶Ж¬·ўУэөДИұПЭЎЈКөСйҪб№ыМбКҫХвЦЦИұПЭКЗУЙУЪИұЙЩmiRNAsјУ№Ө¶шФміЙөДЎЈ¶аКэөДЦІОпmiRNAsФЪДіР©МШ¶ЁЧйЦҜЦРёЯЛ®ЖҪұнҙпТІМбКҫЛыГЗҝЙДЬІОУлБЛЦІОпЧйЦҜөД·ўУэЎЈ
¶ФТ»Іҝ·ЦmiRNAsөДСРҫҝ·ЦОцМбКҫЈәmiRNAsІОУлЙъГь№эіМЦРТ»ПөБРөДЦШТӘҪшіМЈ¬°ьАЁФзЖЪ·ўУэ(Reinhart 2000)Ј¬Пё°ыФцЦіЈ¬Пё°ыөтНцЈ¬Пё°ыЛАНц(Brennecke 2003Ј©Ј¬Ц¬·ҫҙъР»ЈЁXu 2003)әНПё°ы·Ц»Ҝ(Kawasaki 2003)ЎЈҙЛНвЈ¬Т»ёцСРҫҝұнГчЈ¬2ёцmiRNAsЛ®ЖҪөДПВҪөәНВэРФБЬ°НПё°ы°ЧСӘІЎЦ®јдөДПФЦшПа№ШЈ¬МбКҫmiRNAsәН°©ЦўЦ®јдҝЙДЬУРЗұФЪөД№ШПө(Calin 2002)ЎЈ
УЙУЪmiRNAsҙжФЪөД№г·әРФәН¶аСщРФЈ¬МбКҫmiRNAsҝЙДЬУР·ЗіЈ№г·ә¶аСщөДЙъОп№ҰДЬЎЈҫЎ№Ь¶ФmiRNAөДСРҫҝ»№ҙҰУЪіхј¶ҪЧ¶ОЈ¬ҫЭНЖІвmiRNAsФЪёЯј¶ХжәЛЙъОпМеДЪ¶Ф»щТтұнҙпөДөчҝШЧчУГҝЙДЬәНЧӘВјТтЧУТ»СщЦШТӘЎЈУРТ»ЦЦҝҙ·ЁКЗЈәmiRNAsҝЙДЬҙъұнФЪТ»ёцРВ·ўПЦөДІгҙОЙПөД»щТтұнҙпөчҝШ·ҪКҪЎЈ
И»¶шЈ¬ҙу¶аКэmiRNAsөД№ҰДЬИФИ»КЗёцГХЎЈ
miRNAөДЧчУГ·ҪКҪ
ЧоФзұ»·ўПЦөДБҪёцmiRNAsЎӘЎӘlin-4 and let-7ұ»ИПОӘКЗНЁ№эІ»НкИ«»ҘІ№ҪбәПөҪДҝұк°РmRNA 3ЎҜ·ЗұаВлЗш¶ЛЈ¬ТФТ»ЦЦОҙЦӘ·ҪКҪУХ·ўө°°ЧЦК·ӯТлТЦЦЖЈ¬Ҫш¶шТЦЦЖө°°ЧЦКәПіЙЈ¬Чи¶ПmRNAөД·ӯТлЎЈ¶аёц№ыУ¬miRNAsТІұ»·ўПЦәНЛыГЗөДДҝұк°РmRNAsөД3ЎҜ·ЗұаВлЗшУРІҝ·ЦН¬ФҙЎЈУЙУЪmiRNAsәНЖдЗұФЪөДДҝұк°РЦ®јдІў·ЗНкИ«»ҘІ№Ј¬ХвК№өГНЁ№эРЕПўС§өД·Ҫ·Ёјш¶ЁmiRNAөДДҝұк°РО»өгұдөГА§ДСЎЈТт¶шТІОЮ·ЁИ·¶ЁmiRNAsөДЧчУГ·ҪКҪКЗКІГҙЈ¬ТФәОЦЦ»ъЦЖУ°ПмmRNAөД·ӯТлЈ¬ТФәОЦЦ·ҪКҪөчҝШ»щТтұнҙпЎЈmiRNAsөДЧчУГДҝұк°РәН»оРФ»ъЦЖТ»ЦұКЗёчөШөДСРҫҝИЛФұөД№ШЧўИИөгЎЈ
ФЪЦІОпЦРДҝЗ°УРТ»ёцmiRNAәН3ёцЗұФЪөДДҝұк°Р»щТтНкИ«»ҘІ№ЈЁХвР©scarecrow »щТтұаВлЗұФЪөДЧӘВјТтЧУЈ©Ј¬ҫЎ№ЬДҝЗ°»№І»ЗеіюХвР©»щТтКЗ·сҫНКЗmiRNAөДДҝұк°РЈ¬ХвИФКЗөЪТ»ҙО·ўПЦmiRNA әНЖдЗұФЪөДДҝұк°РНкИ«»ҘІ№Ј¬ТІМбКҫmiRNAҝЙДЬ°ьә¬әНsiRNAАаЛЖөДЧчУГ·ҪКҪЎЈ
К¶ұр miRNAsөД·Ҫ·Ё
¶аёцСРҫҝРЎЧйІЙУГЙъОп»ҜС§ҪбәПКЗЙъОпРЕПўС§өД·Ҫ·ЁҝӘХ№¶ФmiRNAsөДСРҫҝ№ӨЧчЎЈУЙУЪҫЭНЖІв¶јКЗУЙDicerГёҪөҪвRNAөГөҪөДЈ¬21ЎӘ23ёцјо»щҙуРЎЎўУР5ЎҜ¶ЛБЧЛб»щәН3ЎҜфЗ»щөДRNAЖ¬¶ПЈ¬УРөДКөСйКТІЙУГёДБјөД¶ЁПтҝЛВЎ·Ҫ·ЁАҙЙёСЎҫЯУРПаН¬МШХчөДРЎ·ЦЧУЎӘЎӘЙёСЎТ»¶ЁҙуРЎөДRNA·ЦЧУЈ¬Б¬ҪУөҪ3ЎҜәН5ЎҜөДККЕдЧУЈЁadaptersЈ©Ј¬ДжЧӘВјІўНЁ№эPCRА©ФцЎўСЗҝЛВЎІўІвРтЎЈmiRNAЗ°МеФЪ»щТтЧйЙПөД¶ЁО»әНҫЫАаКЗНЁ№эПт»щТтЧйКэҫЭҝвІйСҜҪшРРЎЈХвёц·Ҫ·ЁУРЦъУЪЕР¶ПmiRNAsКЗ·сКЗmRNAsЎўtRNAsЎўrRNAsөИ·ЦЧУөДҪөҪвІъОпЎЈ
УРөДКөСйКТНЁ№эТ»ЦЦRNA folding program ЎҜmfoldЎҜ АҙЕР¶ПC. elegans әНC. briggsae Ц®јдөДёЯ¶ИұЈКШЗшУтКЗ·сә¬УРЗұФЪөДmiRNAЗ°МеЈ¬И»әуУГNorthern BlotsөД·Ҫ·ЁАҙИ·¶ЁХвР©miRNAsКЗ·сХжөДұнҙпБЛЎЈ
ҫЎ№ЬУРКэ°ЩёцmiRNAsНЁ№эЙъ»Ҝ»тХЯКЗЙъОпРЕПўС§өД·Ҫ·Ёұ»јшұріцАҙЈ¬ТСҫӯјшұріцАҙөДmiRNAsЦ»І»№эКЗІЧәЈТ»ЛЪЈ¬УЙУЪәЬ¶аТСҫӯјшұріцАҙөДmiRNAsКЗҙУөҘёцҝЛВЎЦРјшұріцАҙөДЈ¬ЛщТФҝЙТФјЩЙи»№УРәЬ¶аmiRNAsФЪ·ЦАләНјш¶Ё№эіМЦРұ»Ў°В©өфЎұБЛЈ¬ІвРт№ӨЧч»№Ф¶Ф¶І»№»ЎЈ
miRNAәНsiRNAөД№ШПө
miRNAәНsiRNAЦ®јдөД№ШПөБоИЛГФ»уЎЈҙУұнГжЙПЛөЈ¬Т»ёцКЗ·ЗұаВлөДөҘБҙРЎ·ЦЧУRNAЈ¬ФЪҪш»ҜЙПёЯ¶ИұЈКШЈ¬НЁ№э·ӯТлТЦЦЖөчҝШ»щТтұнҙп¶шІ»У°ПмЧӘВјұҫөДОИ¶ЁРФЈ»БнТ»ёцКЗХл¶ФұаВлЗшөДЛ«БҙРЎ·ЦЧУRNAЈ¬ГҝёцЧӘВјұҫ¶јҝЙДЬУРәЬ¶аёцsiRNAsЈ¬КЗНЁ№эҪөҪвДҝұк°РЈ¬ФЪЧӘВјәуөчҝШ»щТтұнҙпЎЈУЙУЪГҝёцmRNAДЈ°жҝЙДЬІъЙъәЬ¶аёцsiRNAsЈ¬ТӘёшГҝёцsiRNA¶ЁТ»ёц»щТтөДГыЧЦҫНәЬА§ДСЎЈmiRNAКЗҪш»ҜҪшіМЦРёЯ¶ИұЈКШөДЈ¬ТтҙЛёшЦұПтН¬ФҙОпТ»ёцН¬СщөДГыЧЦҝЙДЬУРЦъУЪБЛҪвЛыГЗөД№ҰДЬЈ¬¶шёшБнТ»ёцОпЦЦЦРТ»¶ООЮ№ШөДРтБРТ»ёцН¬СщөДГыЧЦҫНИЭТЧФміЙ»мВТЎЈ
И»¶шЈ¬ҫЭНЖІвmiRNAsНЁіЈКЗУЙҪПҙуөДЈЁ7090 ntЈ©өДҫҘ»·Ҫб№№ЈЁ·ўјРҪб№№Ј©З°МеҫӯDicerГёЗРёоөГөҪөДЈ¬¶шDicerН¬СщёәФрҪ«іӨЛ«БҙRNAЗРёоОӘsiRNAЈ¬¶шЗТ¶юХЯөДіӨ¶ИТІІоІ»¶аЈ¬Н¬СщУРөчҝШ»щТтұнҙп№ҰДЬЎЈТт¶шХвБҪАаРЎ·ЦЧУRNAЦ®јдөД№ШПөёсНвБоИЛ№ШЧўЎЈ
БҪёц№гОӘИЛЦӘөДmiRNAЎӘЎӘФЪПЯіжЦРКЧҙО·ўПЦөДlin-4 әНlet-7Ј¬ҝЙТФНЁ№эІҝ·Ц»ҘІ№ҪбәПөҪДҝөДmRNA°РөД3ЎҜ·ЗұаВлЗш(3ЎҜUTRs)Ј¬НЁ№эТ»ЦЦОҙЦӘ·ҪКҪУХ·ўө°°ЧЦК·ӯТлТЦЦЖҙУ¶шТЦЦЖө°°ЧЦКәПіЙЎЈХвЦЦҪбәПІўІ»УХөјmRNA°РөДҪөҪвЈ¬ҫНКЗЛөЧчОӘ·ӯТлТЦЦЖЧУұҫЙнІ»У°Пм¶ФУҰmRNAөД·б¶ИЈ¬ЖдФӯТтҫЭНЖІвКЗУЙУЪmiRNAәНҪбәПО»өгЦ®јдІ»НкИ«»ҘІ№ЎЈХвҫНЗшұрУЪsiRNAөДҪйөјөДmRNAөДҪөҪвЎЈө«КЗЖдЛыТ»Р©miRNAsҝЙДЬТФАаЛЖsiRNAөД·ҪКҪҪйөјДҝөДRNAөДҪөҪвЎЈКөСйұнГчТэИләНlet-7ДҝөДmRNA°РНкИ«»ҘІ№өДmiRNA»бУХөјmRNA°РөДҪөҪвЎЈ»№УРКөСйҪб№ыұнГчТ»Р©miRNAЈ¬°ьАЁФЪЦІОпЦР·ўПЦөДScarecrow miRNAЈ¬ДЬҪбәПНкИ«»ҘІ№өДmRNAБҙҙУ¶шҪөҪвmRNAРтБРЈ¬ТЦЦЖө°°ЧәПіЙЎЈХвМбКҫmiRNAsҝЙТФәНsiRNAsТ»СщЧчУГЈ¬ХвБҪЦЦРЎ·ЦЧУRNAЧчУГНЁВ·ҝЙДЬУРЦШөюөДІҝ·ЦЎЈХвЦЦЦШөюН¬СщМбКҫsiRNAsҝЙДЬТІУРәНmiRNAsН¬СщөД№ҰДЬЎЈ
Т»ёцәЬУРИӨөДКөСйЦӨКөХвёц№ЫөгЈәDoenchәНН¬КВМфСЎТ»ёцТСЦӘФЪМеДЪҝЙТФУРР§К№CXCR4»щТтіБД¬өДsiRNAЈ¬И»әуФЪУ«№вЛШГёұЁёж»щТтөД3ЎҜ¶ЛІеИл¶ФУҰөДCXCR4ҪбәПО»өгЎӘЎӘЖдЦРТ»ёцҝҪұҙКЗІеИлТ»ёцНкИ«ЖҘЕдөДCXCR4ҪбәПО»өгЈ¬БнТ»ёцҝҪұҙІеИл4ёцЦ»УР3ЎҜәН5ЎҜ¶ЛЖҘЕдЈ¬¶шЦРјдІ»Н¬өДCXCR4ҪбәПО»өгЈ¬ХвСщСЎ¶ЁөДsiRNAҫНІ»ДЬНкИ«ҪбәПөҪХвёцҪбәПО»өгЎӘЎӘЦРјдРОіЙТ»ёцН»ЖрөДІ»ЖҘЕдөД»·ЎЈҪ«ХвБҪёцҝҪұҙЧӘИлHelaПё°ыІўУГsiRNAУХөј»щТтіБД¬ЎЈҪб№ыәЬУРИӨЎӘЎӘБҪёцКөСй¶јВјөГУ«№вЛШГё»оРФПВҪөБЛі¬№э10ұ¶Ј¬RT-PCRәНNorthern·ЦОцЦӨКөЈ¬өЪТ»ёцКөСйөДУ«№вЛШГёЧӘВјұҫПВҪөБЛі¬№э10ұ¶Ј¬ХвХэКЗХэіЈөДsiRNAҪйөјөДRNAi·ҙУҰЈ¬Дҝұк°РmRNAҪөҪвөјЦВұнҙпЛ®ЖҪөДПВҪөЈ¬¶шөЪ¶юёцКөСйЦРУ«№вЛШГёЧӘВјұҫҪцҪцПВҪө1.2ұ¶Ј¬ХвЦЦДҝөД»щТтұнҙпЛ®ЖҪПВҝҙЖрАҙПуФҙУЪmiRNAҪйөјөД·ӯТлТЦЦЖҪөЈ¬¶шІ»КЗsiRNAҪйөјөДУ°ПмmRNAөДОИ¶ЁРФөјЦВЎЈКөСйұнГчЈәsiRNAҝЙДЬТФmiRNAөД·ҪКҪЧчУГУЪmRNAЎЈКөСйИЛФұ»№ҪшРРБЛБнТ»ёцКөСйЈәёДұдөЪ¶юёцКөСйЦРөДІ»ЖҘЕд»·өДјо»щРтБРҝҙЖрАҙІ»У°ПмТЦЦЖР§№ыЈ¬ө«КЗsiRNAәНұЁёж»щТтЙПөДҪбәПО»өгөДЖҘЕдіМ¶ИФҪёЯТЦЦЖР§№ыФҪәГЈ¬ФцјУsiRNAөДБҝЈ¬ТЦЦЖР§№ыФҪәГЎӘЎӘХвТ»өгәНsiRNAТЦЦЖөДЗйҝцТ»СщЎӘЎӘОЁТ»І»Н¬өДКЗЈәНкИ«ЖҘЕдөДҪбәПО»өгЈЁsiRNAЧчУГ·ҪКҪЈ©ҝЙТФөҘ¶АЖрЧчУГ¶шПа»ҘІ»У°ПмЈ¬¶шФцјУІ»НкИ«Ед¶ФөДҪбәПО»өгЈЁЧўТвФЪөЪ¶юёцКөСйЦРУГБЛ4ёцCXCR4ҪбәПО»өгЈ©өДёцКэ¶Ф·ӯТлТЦЦЖУРПФЦшөДјУіЛЧчУГЎЈ
ФЪІёИй¶ҜОпПё°ыЦР»№Г»УРХТөҪДЪФҙөДsiRNAЈ¬НвФҙөДsiRNAҪйөјөДRNAiЧчУГХэКЗТ»ЦЦөЦУщ»ъЦЖЎЈ¶шmiRNAsФт№г·әҙжФЪУЪІёИй¶ҜОпПё°ыЦРЈ¬ҙУАнВЫЙПНЖІвҝЙДЬІОУл¶аЦЦөчҝШЧчУГЎЈХвБҪЦЦРЎ¶«ОчөДЧчУГ»ъЦЖәНПа»Ҙ№ШПөөДұҫЦКҫНПФөГёьјУЖЛЛ·ГФАлЎЈИзәОФЪКөСйЦРХэИ·јш¶ЁsiRNAәНmiRNAЈ¬ЙхЦБКЗЖдЛыөДРЎ·ЦЧУRNA¶јіЙОӘТ»ёцЦөөГ№ШЧўөДОКМвЎЈ
ОҙАҙТӘҪвҫцөДОКМв
miRNAsФЪ¶аёцОпЦЦЦР№г·әұ»·ўПЦЈ¬¶шЗТФЪҪш»ҜЙПёЯ¶ИұЈКШЎЈХвР©Ў°РЎНжТв¶щЎұБфёшОТГЗТ»ҙу¶СГХНЕЈәmiRNAөДИ·ЗР№ҰДЬКЗКІГҙЈҝЛьөДДҝұк°РКЗКІГҙЈҝЧчУГ»ъЦЖКЗКІГҙЈҝТІРнРиТӘ¶ФЦІОп»тХЯПЯіжөД»щТтЧйҪшРРmiRNAsН»ұдЦкөДЙёСЎЈ¬ФЪ№ыУ¬ЦРҝЙТФУГtargeted-disruptionИұК§miRNAРтБРЎЈ¶ФmiRNAН»ұдЦк°йЛжөДұнРНИұК§ҪшРРСРҫҝЈ¬УРЦъУЪҪвКНmiRNAsөД№ҰДЬЎЈ
ХэИзPhillip ZamoreЛөөДЈәЎ°Из№ыmiRNAsФЪҪш»ҜөДҪшіМЦРИзҙЛёЯ¶ИұЈКШ¶шГ»УРИОәОКөјК№ҰДЬЈ¬ДЗХжКЗҙуЧФИ»ДГҝЖСРИЛФұҝӘдМЎӘЎӘ¶шЗТКЗТ»ёцІРҝбөДНжРҰЎұЎЈ
СРҫҝmiRNAөДРВ№ӨҫЯ
ЛжЧЕРЎ·ЦЧУRNAИХТжКХКЬөҪСРҫҝИЛФұөДЦШКУЈ¬әЬ¶аСРҫҝРЎ·ЦЧУRNAөДРВ·Ҫ·ЁІ»¶ПНЖіцЎЈ
РЎ·ЦЧУRNAөД·ЦАл
УЙУЪРЎ·ЦЧУRNAҝЙДЬІОУл·Ц»ҜЎў·ўУэЎўЧйЦҜЙъіӨЎўЦ¬·ҫҙъР»өИЙъАн№эіМЈ¬ФЪІ»Н¬өДЧйЦҜәН·ўУэҪЧ¶ОөДұнҙпЛ®ЖҪУРЛщІ»Н¬Ј¬ҪшТ»ІҪБЛҪвРЎ·ЦЧУRNAөДЙъОп№ҰДЬРиТӘИ·¶ЁЖдФЪёчЦЦЙъОпСщЖ·ЦРөДұнҙпЛ®ЖҪЈ¬Тт¶шРиТӘТ»ЦЦҫ«И·өД¶ЁБҝҙҝ»Ҝ·Ҫ·ЁЈ¬ҙУ¶шөГөҪҝЙРЕөДКэҫЭЎЈ
ПЦРРөДRNAҙҝ»Ҝ·Ҫ·Ё°ьАЁУР»ъИЬјБійМб+ТТҙјіБөнЈ¬»тХЯКЗІЙУГёьјУ·ҪұгҝмҪЭөД№иҪәДӨАлРДЦщөД·Ҫ·ЁАҙҙҝ»ҜRNAЎЈУЙУЪ№иҪәДӨАлРДЦщНЁіЈЦ»ё»јҜҪПҙу·ЦЧУөДRNAЈЁ200ntТФЙПЈ©Ј¬РЎ·ЦЧУRNAНщНщұ»МФМӯөфЈ¬Тт¶шІ»ККУГУЪРЎ·ЦЧУRNAөД·ЦАлҙҝ»ҜЎЈУР»ъИЬјБійМбДЬ№»ҪПәГөДұЈБфРЎ·ЦЧУRNAЈ¬ө«КЗәујМөДіБөнІҪЦиұИҪП·СКұ·СБҰЎЈmirVana miRNA Isolation KitКЗІЙУГІЈБ§ПЛО¬ВЛДӨАлРДЦщ(glass fiber filterЈ¬GFF)Ј¬јИДЬ№»УРР§ё»јҜ10merТФЙПөДRNA·ЦЧУЈ¬УЦДЬ№»јжұёАлРДЦщҝмЛЩАлРДҙҝ»ҜөДУЕөгЈ¬КЗТ»ёцІ»ҙнөДСЎФсЎЈ¶ФУЪМШұрПЎУРөД·ЦЧУЈ¬УЙУЪРиТӘ·ЦАлҙуБҝRNA¶шөјЦВёЯұіҫ°¶шҪөөНБйГф¶ИЈ¬»№ҝЙТФҪшТ»ІҪё»јҜ10merөҪ200bpөДРЎ·ЦЧУRNAАҙМбёЯБйГф¶ИЎЈ
РЎ·ЦЧУRNAМҪХлөДЦЖұё
·Ҫ·ЁЖдКөәЬјтөҘЈәЦ»РиТӘЧјұёДҝөД»щТтөДТ»РЎ¶О№СәЛЬХЛбРтБРЈ¬3ЎҜ¶ЛБнНвФцјУ8ёцәНT7Жф¶ҜЧУ»ҘІ№өДјо»щЈ¬Ҫ«Хв¶О№СәЛЬХЛбәНT7Жф¶ҜЧУТэОпНЛ»рЈ¬УГKlenowҙуЖ¬¶ПІ№ЖлөГөҪЛ«БҙөДЧӘВјДЈ°жЈ¬И»әуУГT7 RNAҫЫәПГёЎўrNTPәНұкјЗОп»мәПЈ¬МеНвЧӘВјөГөҪұкјЗөДРЎ·ЦЧУRNAМҪХлЎЈХвЦЦ·Ҫ·ЁҝЙТФҝмЛЩЦЖұёёчЦЦұкјЗЈЁН¬О»ЛШЎў·З·ЕЙдРФұкјЗҫщҝЙЈ©өДРЎУЪ100ntөДРЎ·ЦЧУRNAМҪХлЈ¬ККУГУЪ°ьАЁRPAsЈ¬Northerns әНФӯО»ФУҪ»өИёчЦЦ·Ҫ·ЁјмІвРЎ·ЦЧУәЛИКRNA( small nuclear RNA,snRNA)Ј¬small interfering RNA (siRNA),Ј¬micro RNA (miRNA)әН mRNAЎЈ·З·ЕЙдРФұкјЗөДМҪХл»№ҝЙТФУГУЪФӯО»ФУҪ»СРҫҝmiRNA»тХЯmRNAФЪЧйЦҜЦРөД·ЦІјЎЈ
РЎ·ЦЧУRNAөДјмІв
УЙУЪРЎ·ЦЧУRNAКЗТ»АаәЬРЎөД·ЦЧУЈ¬Іҝ·ЦРЎ·ЦЧУRNAұнҙпЛ®ЖҪҝЙДЬәЬөНЈ¬Тт¶шРиТӘј«ОӘБйГф¶ш¶ЁБҝөД·ЦОц№ӨҫЯЎЈУЙУЪЖд·ЦЧУәЬРЎЈ¬УГRT-PCRөД·Ҫ·ЁАҙ¶ЁБҝСРҫҝ·ЗіЈА§ДСЈ¬ДҝЗ°¶аКэСРҫҝИЛФұІЙУГNorthern BlotsЎӘЎӘТ»ЦЦјјКхёҙФУ¶ш·СБҰөД·Ҫ·ЁАҙјмІвРЎ·ЦЧУRNAөДҙжФЪЎЈҙ«НіөДNorthern BlotөД·Ҫ·ЁКЗКЗУГМҪХлјмІв№МПаЦ§іЦОпЈЁДӨЈ©ЙПөДДҝұк·ЦЧУЈ¬УЙУЪУГМҪХлјмІвТәПаЦРөДДҝұк·ЦЧУФ¶ұИјмІв№МПаЦРөДДҝұкёьОӘБйГфЈ¬ЙъОпНЁФЪХвАпОӘҙујТНЖјцТ»ЦЦ»щУЪәЛГёұЈ»Ө·ЦОц·Ҫ·ЁёДҪшөДРВ·Ҫ·ЁЎӘЎӘҪ«Н¬О»ЛШұкјЗәГөДРЎ·ЦЧУRNAМҪХләНҙэјмІвСщЖ·»мәПФУҪ»Ј¬ОҙФУҪ»өДRNAәН¶аУаөДМҪХлУГөҘБҙәЛЛбГёПы»ҜЈ¬И»әуК№әЛЛбГёК§»оЈ¬Іўҙҝ»ҜФУҪ»өДRNA·ЦЧУЈ¬ЧоәуНЁ№эұдРФҪәөзУҫ·ЕЙдЧФПФУ°јмІвҪб№ыЎЈХвёц»щУЪТәПаФУҪ»өДРВ·Ҫ·ЁІ»ө«ІЩЧчјтөҘ¶шҝмЛЩЈ¬¶шЗТБйГф¶Иј«ёЯЎӘЎӘҝЙТФ°л¶ЁБҝјмІвЙЩЦБ10ngЧЬRNAДЈ°жЦРөДРЎ·ЦЧУRNAЈ¬»тХЯЛөЈ¬ҝЙТФјмІвattomole (10-18 mol)ј¶ұрөД°РДҝұкЈЎБйГф¶ИКЗNorthern BlotөД100ұ¶ЎЈіэҙЛЦ®НвЈ¬СРҫҝИЛФұ»№ҝЙТФФЪН¬Т»ёцСщЖ·ЦРН¬КұјмІв¶аёцРЎ·ЦЧУRNAәНіӨөДRNAДЈ°жЎЈУҰёГЛөЈ¬ХвёцБй»оЗЙГоөДЙијЖҝЙТФОӘҙУКВРЎ·ЦЧУRNAКөСйөДСРҫҝИЛФұҙшАҙІ»ЙЩ·ҪұгЎЈ
ЧЬ¶шСФЦ®Ј¬ОЮВЫКЗsiRNA, miRNA, snRNA»№КЗЖдЛыөДРЎ¶«ОчЈ¬РЎ·ЦЧУRNAСРҫҝөДІ»¶ПЙоИлҪ«°пЦъОТГЗҪТКҫёь¶аЙъГьөД°ВГШЎЈ
ҪсИХ¶ҜМ¬ | ИЛІЕКРіЎ | РВјјКхЧЁАё | ЦР№ъҝЖС§ИЛ | ФЖХ№МЁ | BioHot | ФЖҪІМГЦұІҘ | »бХ№ЦРРД | МШјЫЧЁАё | јјКхҝмС¶ | Гв·СКФУГ
°жИЁЛщУР ЙъОпНЁ
Copyright© eBiotrade.com, All Rights Reserved
БӘПөРЕПдЈә
ФБICPұё09063491әЕ



