
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
m6A调节果蝇性别分化和神经功能 | 案例解析
【字体: 大 中 小 】 时间:2018年05月08日 来源:联川生物
编辑推荐:
听听专家解析m6A研究的具体案例――m6A调节果蝇性别分化和神经功能。
标题:m6A modulates neuronal functions and sex determination in Drosophila
m6A调节果蝇性别分化和神经功能
发表时间:2016年12月
发表期刊:Nature
IF:40.1
机构:德国美因茨大学
1、前言
RNA修饰在表观转录调控中十分重要,其中m6A修饰在许多哺乳动物中广泛存在。许多真核生物中都存在m6A修饰,包括人、小鼠、斑马鱼甚至酵母。许多研究表明大量的m6A修饰落在了RRACH motif(R, purine; H, non-guanine base)内,另外终止密码子和3’ UTR区域也有大量的m6A修饰。RNA甲基化修饰参与许多转录后调控,包括mRNA前体剪切,mRNA降解和翻译,其中的“reader”酶YTH家族起到十分重要的作用。RNA甲基化转移酶(methyltransferase)能够催化腺苷酸N6位发生甲基化,包括METTL3、METTL14、WTAP、Vir等。而去甲基化酶包括FTO和ALKBH5。
已知在小鼠胚胎干细胞中将METTL3敲除(knockout)后能够阻止其发生分化,MELLT3基因功能缺失对小鼠胚胎干细胞的发育具有致死做用。同样在果蝇中,METTL3的同源蛋白Ime4被敲除后也具有亚致死作用,由于NOTCH信号通路受损从而影响果蝇的生育能力。在模式植物拟南芥中,METTL3同源蛋白Ath-MTA被敲除后,其发育也会受到很大影响。另外,Ime4基因对酵母的减数分裂也有很重要的作用。种种迹象表明,m6A修饰对生物早期性器官发育以及早期的胚胎发育至关重要。近几年的结构生物学研究表明,METTL3和METTL14会形成杂络物行使催化作用,但是生物体内的作用机制至今仍然未知。参与RNA甲基化的酶包括:
|
类别 |
基因 |
功能 |
|
Writers |
METTL3, METTL14, WTAP, KIAA1492 |
METTL3和METTL14形成杂络物,在体内和体外催化RNA发生m6A甲基化修饰。而WTAP(Fl(2)d)和KIAA1492(Vir)以及其他factors也杂络物的重要组成部分。这些Writers都有consensus motif |
|
Erasers |
FTO, ALKBH5以及其他homologies |
去甲基化酶,介导m6A去甲基化修饰 |
|
Readers |
YTHDF1、YTHDF2、YTHDF3以及其他同源家族蛋白 |
识别RNA甲基化修饰的信息,并参与下游RNA的翻译、降解等过程。这些YTHDF家族的蛋白普遍都含有YTH domain |
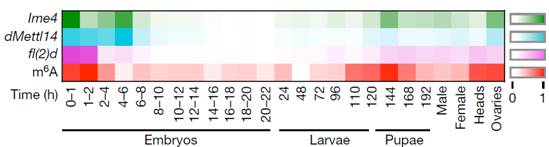
昆虫整个生命周期包括胚胎期、幼虫期(一龄幼虫→三龄幼虫)、蛹期以及成虫期。作者先检测了野生型果蝇各个时期的m6A整体的甲基化水平,使用质谱法对Ime4、dMettl14和fl(2)d三种蛋白相对表达水平做了一个热图。从热图中我们可以发现在胚胎早期尤其是2小时后m6A水平处于一个很高的水平,之后开始下降。在三龄幼虫期,m6A水平开始上升,在蛹期又达到了一个高峰后逐渐下降。果蝇成虫期的头部以及卵巢中m6A水平也一直维持相对较高水平。
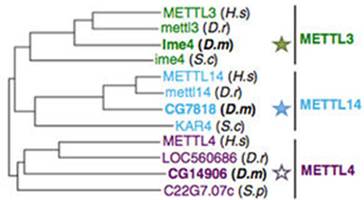
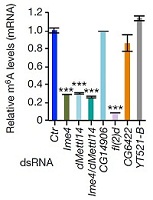
系统进化树分析表明,果蝇的METTL3同源蛋白Ime4有两种密切相关的蛋白CG7818和CG14906。在果蝇的SR2+细胞(embryonic-derived Schneider)中敲低Ime4和CG7818后,m6A水平下降70%而敲低CG14906却没有任何影响。由于CG7818与METTL14结构上具有很大的保守性和相似性,作者将CG7818重新命名为dMettl14。同样fl(2)d和Vir基因与WTAP和KIAA1429高度同源,敲低后对果蝇的m6A水平也会产生较大的影响。
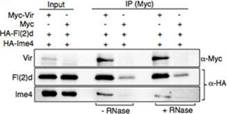
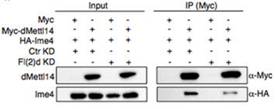
在哺乳动物尤其是人体内中,RNA甲基化转移酶METTL3和METTL14会形成杂络物,而WTAP和KIAA1429也是重要的组成部分。在果蝇甲基化转移酶中同时发现了Vir、fl(2)d和Ime4。作者对果蝇的fl(2)d敲低后,Ime4和dMettl14的interaction显著降低。
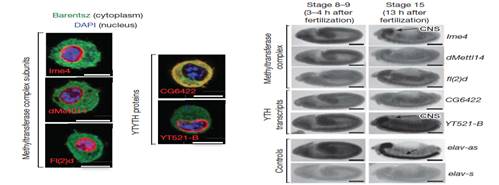
基因定位实验表明参与m6A甲基化的酶都位于细胞核内且都在胚胎早期发育阶段处于高表达水平。另外胚胎晚期,这些酶在神经外胚层也有较高的转录水平。
3.YT521-B介导m6A调控mRNA可变剪切
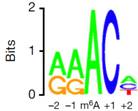
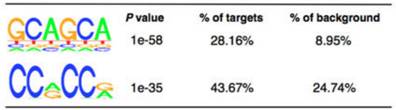
作者使用MeRIP-seq,从812个基因中鉴定了到了1120个m6A的peak。其中92%的peak都含有RRACH motif。此外,其他类型的序列或motifs也含有较高的m6A水平,这表明这些motifs可能也对甲基化转移酶具有特异性。

此外,与其他真核生物类似,在果蝇体内start codon和stop codon区域存在很高的m6A甲基化修饰。
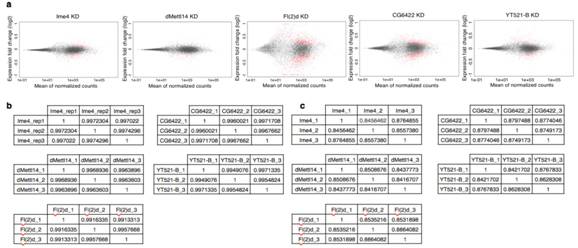
接下来,作者针对SR2+细胞系中与m6A相关的各种酶敲低后进行了转录组测序,这些酶包括Ime4、CG6422、dMellt14、YT521-B以及Fl(2)d。从上图a中我们可以明显看到,MA-plot显示不同敲低条件下,SR2+细胞系中的基因表达发生了显著的差异(p<0.05)。图b表示不同敲除条件下三个生物学重复基因表达的spearman相关系数表。图c则是剪切模式的spearman相关系数表。
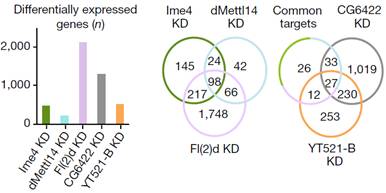
其中作者发现Fl(2)d敲低后效果最为明显,超过2000个基因表达显著差异。敲低Ime4和dMettl14后,通过上面的韦恩图可以发现SR2+中差异表达基因数量小于Fl(2)d(milder effects)。
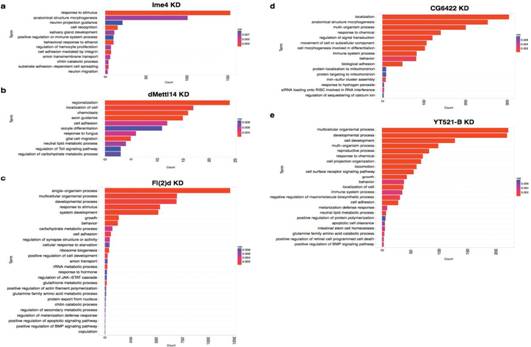
GO功能富集分析表明,这些差异表达的基因涉及各种功能如阴离子转运、细胞粘附等。尽管SR2+细胞系并不是直接起源于神经系统,但是许多差异基因都富集到了neuronal functions上,如axon guidance和synapse activity。

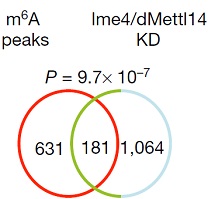
通过表达量盒形图发现,基因较低组的细胞中整体表达水平显著高于control组。接下来,对SR2+细胞系中的Ime4和dMettl14这2个基因进行双敲低(double knockdown)。韦恩图显示至少15%的基因都有m6A的peak。
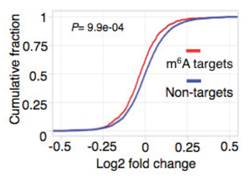
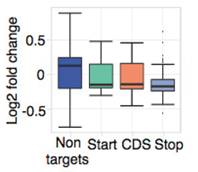
Ime4和dMettl14双敲后,m6A甲基化水平仍然发生了轻微但是仍然显著的差异。这个结果表明每个转录本可能拥有独立的m6A peak位点。
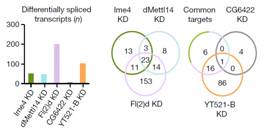
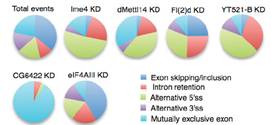
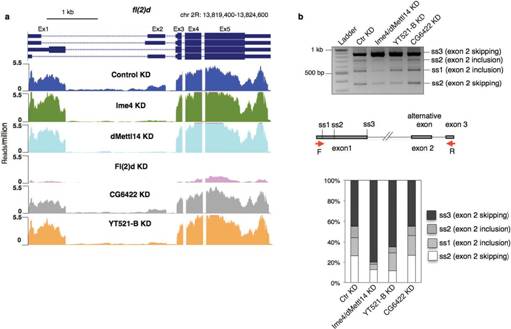
文章还发现在基因剪切事件上,双敲的结果与2种酶单独敲的结果低还是存在显著差异的。在任意一种酶的敲低实验中,fl(2)d这个基因都行使十分重要的作用。通常情况下,不同酶敲低都会对基因5’的剪切以及内含子保留等行为产生影响,这种情况同样也存在于人细胞系中。

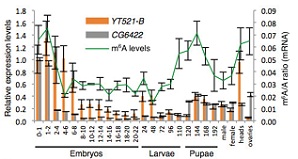
YTH家族蛋白是一种十分重要的reader酶。果蝇中,CG6422和YT521-B属于YTH家族的同源蛋白。蛋白细胞定位实验表明CG6422位于细胞质中,受精后在胚胎期2小时达到最大值,接下来持续下降并且在幼虫期和蛹期一直维持较低的水平。然而,蛋白细胞定位实验表明YT521-B位于细胞核中,并且在胚胎的神经系统已经成虫的大脑中处于较高的转录水平。
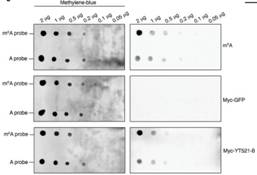
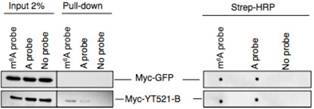
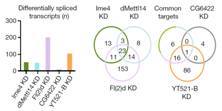
使用点印记法(dot-plot)和pull down实验表明,YT521-B能够与RNA上的m6A位点相结合。转录组测序表明,敲低CG6422后,差异基因RNA剪切事件几乎影响不大。但是敲低YT521-B后,转录组结果表明有103个基因的剪切事件差异显著。韦恩图显示YT521-B与common target以及CG6422交集不到30%。所以这些证据都表明YT521-B在pre-mRNA剪切上行使极其重要的功能。
4.m6A甲基转移酶对果蝇的行为调控

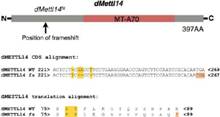
为了寻找m6A相关酶的潜在功能,作者分别构建了Ime4和dMellt14敲除的果蝇。

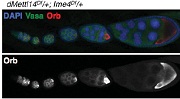
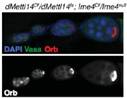
其中针对Ime4这个基因使用两种方法敲除方法,Ime4Null(CDS全部敲除)和Ime4Cat(C端保留催化结构域)。文章发现无论是纯合果蝇还是发生杂合突变的果蝇,幼虫都能羽化存活至成年期。卵巢中没有发现任何encapsulation defects。

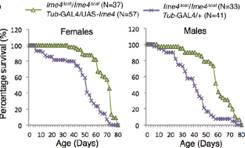
但是突变型果蝇寿命变短,飞行和爬行受阻,无法合翅。尽管突变型果蝇能够勉强合翅伸缩翅膀,但是很难将翅膀合拢在腹部和胸部的背面。另外dMettl14基因突变型果蝇虽然翅发育正常,但是行为能力却已经缺失。为了验证Ime4和dMettl14是否在果蝇体内能够相互补偿,作者构建了双突变果蝇和单基因突变果蝇。结果表明,这2个基因功能相似调控的通路也相似,可能调控common targets。此外,Ime4似乎在功能上比dMettl14更占优势,大部分催化反应需要甲基化转移酶有特定的催化结构域。
小贴士:果蝇属于昆虫纲双翅目,只有一对翅膀,另一对翅膀已经进化成平衡棒,用于辅助平衡飞行。通常翅膀会合拢在胸部和腹部的背面。
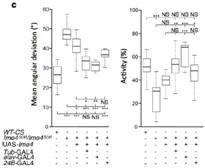

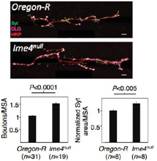
行为实验表明Ime4突变型果蝇爬行速度显著低于野生型果蝇,定向能力(Orientation)也出现问题。种种证据表明m6A修饰通过调控神经系统,从而严重影响果蝇的行为。为了通过表型行为探究深层的神经缺陷和分子机制,作者在Ime4突变型果蝇幼虫中进行了肌肉神经接头测试(NMJ)。上图显示NMJ在Ime4突变型幼虫内树突生长是野生型的1.5倍。结果表明Ime4通过控制NMJ的树突生长来调节果蝇幼虫的爬行行为。
为了鉴定果蝇行为的靶基因,作者对二日龄雌果蝇成虫的头部进行了解剖并进行转录组测序。通过转录组结果分析,作者在Ime4突变型中鉴定了1600多个差异基因在基因剪切模式上发生改变。这些差异基因绝大部分都是控制果蝇行为的。接下来作者又对SR2+细胞系原有的MeRIP数据与转录组数据进行比对,发现大部分调控果蝇行为的基因都发生了m6A甲基化修饰。作者推测,甲基化转移酶缺陷的情况下,可能有不止一个基因调控果蝇的行为。
5.甲基化修饰m6A调控果蝇性别分化
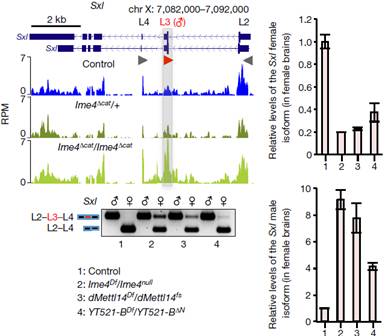
在果蝇体内,sxl基因这个开关激活与否由上游的剂量补偿效应来调控控制果蝇的性别分化,果蝇性别分化机制详见下面小贴士部分。文章内,作者通过实验发现Ime4对Sxl影响很大。Sxl基因在雄虫和雌虫中同时存在,只不过雄虫Sxl基因有一个额外插入的外显子。在突变体雄性果蝇基本不受Ime4的影响,而两种突变型雌性果蝇在基因型上都表现出与雄性果蝇共有的外显子插入以及雌雄果蝇独有的剪切模式。无论是雌虫还是雄虫,Sxl不同转录本下降水平不是特别明显,这表明m6A可能在大脑中特异性富集。
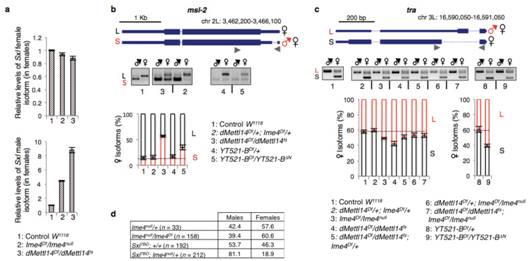
Sxl基因下游调控的2个基因tra和msl也发生了显著变化。这表明甲基化转移酶可能会影响Sxl基因的pre-mRNA的剪切,从而影响果蝇性别分化和剂量补偿效应。为了验证这个猜想,作者使用Ime4杂合突变的雌虫与Sxl杂合突变的雄虫进行杂交,统计后代存活率发现后代雄虫不受影响,Ime4与Sxl缺失的雌性大幅度下降。所以结果表明,Ime4破坏了果蝇体内正常的剂量补偿效应,并通过调控Sxl影响雌雄果蝇的存活。

小贴士:果蝇(左)和家蝇(右)在控制性别分化上采用完全不同的机制。2017年Science刊登论文首次鉴定了家蝇雄性决定基因Mdmd。果蝇通常采用补偿剂量机制(dosage compensation),即当上游的X:A的常染色体和性染色体比例决定下游的Sxl是否会被激活。如果Sxl基因受到影响,无论上游的X:A染色体比例如何,果蝇都会发育成雄性成虫。相反,如果Sxl一直处于激活状态,则果蝇会发育成雌性成虫。
6.YT521-B缺失与m6A表型之间的关系
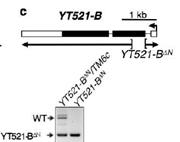
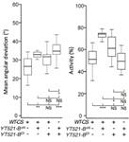
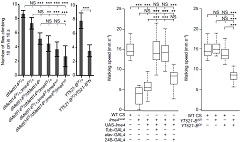
鉴于YT521-B能够特异性识别m6A甲基化位点,从而影响SR2+细胞中绝大部分由m6A介导的RNA剪切事件,作者对各种参与RNA甲基化的酶进行了敲除(knockout)实验。YT521-B缺失能够引起不同转录本表达异常,而YT521-B突变果蝇虽然能够存活至成虫期,但是飞行等行为受到了很大的影响。

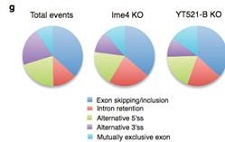
将YT521-B突变雌虫与Ime4突变雌虫进行转录组测序,发现各自有243和397个基因存在剪切模式的差异。5’端可变剪切位点特异性不高,内含子部分保留仍然较多。显然,YT521-B突变也能影响male-specific Sxl,从而影响下游的tra和msl-2,继而使得雌虫体内female-specific Sxl转录水平降低。以上结果表明m6A甲基转移酶是通过YT521-B调控果蝇行为和性别分化。
7.Nito是一种全新的参与m6A修饰的酶
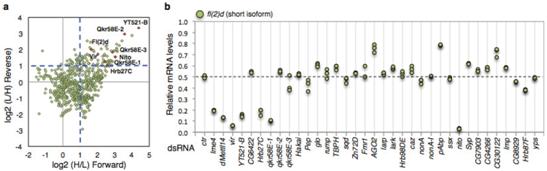
为了探究YT521-B的调控机制,作者利用SILAC技术,对SR2+细胞培养过程中的氨基酸进行同位素标记,结合质谱技术和免疫共沉淀对SR2+细胞中和YT521-B免疫结合的蛋白进行定量分析。几乎一半(n=30)与预测的结合蛋白相符合。为了验证其中是否存在m6A介导的剪切事件,我们分别对这30个蛋白进行了depleted,然后评估fl(2)d对剪切事件的影响。其中蛋白Hrb27C、Qkr58E-1和Nito与fl(2)d控制的剪切现象相符合。
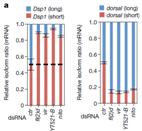
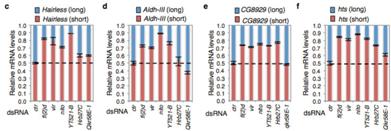
分析了6个额外的m6A介导的剪切事件表明,Hrb27C和Qkr58E-1仅仅调控一个蛋白亚基,而Nito的缺失会导致基因剪切时间异常,这个异常现象与YT521-B及其蛋白复合物功能缺失后的现象比较相似。

接下来作者使用免疫共沉淀将YT521-B与Hrb27C、Qkr58E-1和Nito三种结合蛋白进行共沉淀探寻蛋白间的相互作用。结果表明,Hrb27C和Qkr58E-1与YT521-B的结合依赖于RNA。
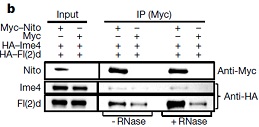


而Nito蛋白与YT521-B蛋白相结合则完全不依赖于RNA。但是Nito蛋白与Ime4以及fl(2)d相结合依赖于RNA。Nito基因的敲低会导致m6A修饰水平的降低。因此Nito缺失对于m6A的水平影响很大。该研究还鉴定到了果蝇m6A甲基转移酶复合体中的新亚基Nito(Spenito)。

