编辑推荐:
2022年1月10日,清华大学药学院谭旭课题组在《Nature Communications》上在线发表了题为“Gain-of-function genetic screening identifies the antiviral function of TMEM120A via STING activation”的研究论文,该研究应用人类全基因组过表达系统,筛选出了新的寨卡病毒的抑制因子TMEM120A,并进一步发现其可以促进STING从ER到ERGIC的转运,从而增强抗病毒先天免疫
2022年1月10日,清华大学药学院谭旭课题组在《Nature Communications》上在线发表了题为“Gain-of-function genetic screening identifies the antiviral function of TMEM120A via STING activation”的研究论文,该研究应用人类全基因组过表达系统,筛选出了新的寨卡病毒的抑制因子TMEM120A,并进一步发现其可以促进STING从ER到ERGIC的转运,从而增强抗病毒先天免疫。
研究背景 早在1947年,人类就从寨卡森林的猴子身上发现并分离出了寨卡病毒(Zika virus)【1】。过去几十年,由于仅在非洲和东南亚呈零星散在性爆发,且感染者仅表现出发热,头疼等较轻症状,并没有引起的足够的重视【2】。自2015年巴西的最近一次爆发,越来越多的证据表明:寨卡病毒感染会导致新生儿小头畸形等神经发育缺陷性疾病 【3, 4】。但是,由于先前的研究不足,到目前为止仍缺乏能有效地对抗寨卡病毒感染的疫苗和治疗策略。
寨卡病毒入侵宿主细胞的过程中,会劫持和降解特定的宿主蛋白,以促进自身的复制;同时,宿主细胞识别病毒的入侵,激活I型干扰素通路等途径,动员抗病毒蛋白以阻止病毒的入侵。全面了解寨卡病毒与宿主蛋白的相互作用,有助于开发新型的治疗方法。2015年以来,多个研究团队运用功能丧失型筛选方法,如CRISPR-Cas9,RNAi 【5-7】,筛选出了多种寨卡病毒相关宿主因子,使我们对寨卡病毒与宿主蛋白的相互作用有了更为清晰的认识。
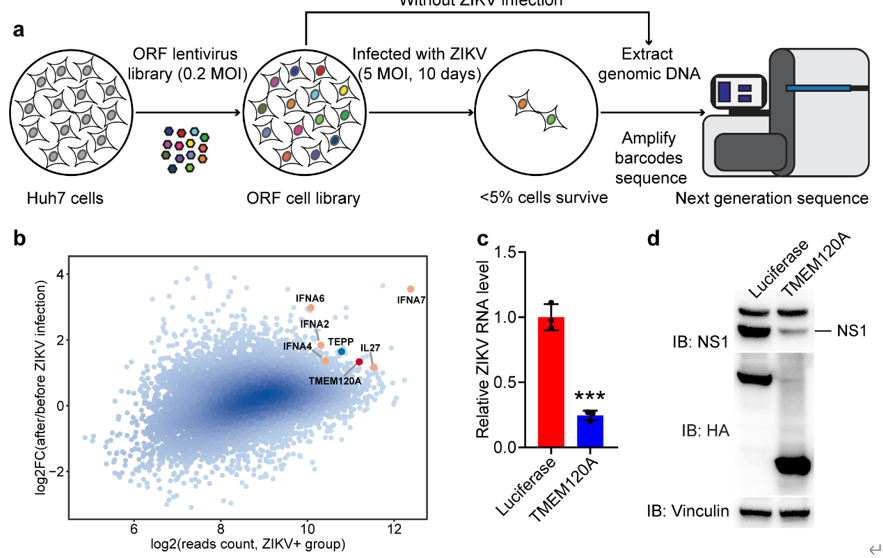
但是,如果使用的细胞系内不表达特定的蛋白,功能丧失型筛选方法就无法筛选出该蛋白,另外,病毒会利用多种宿主蛋白来完成其复制周期中的同一过程,细胞也会动员多种宿主因子对抗病毒的复制周期的同一环节,因此,功能丧失型筛选无法筛选出所有的病毒相关因子。 寨卡病毒抑制因子TMEM120A的发现 考虑到CRISPR-Cas9,RNAi等功能丧失型筛选方法的缺陷,在本研究中,为鉴定新的寨卡病毒宿主因子,作者应用全基因组慢病毒过表达筛选系统,在肝癌细胞系Huh7细胞中进行了功能获得型筛选。该慢病毒过表达系统涵盖了编码人类蛋白的超过16000个开放阅读框(ORF),每个ORF都被一小段特定的随机序列(条形码)所编码。筛选的流程见图1.a。作者将包含ORF的慢病毒质粒转入HEK293T细胞中包被ORF慢病毒文库;慢病毒ORF文库转导Huh7细胞后,用嘌呤霉素筛选转导成功的细胞;用寨卡病毒感染转导成功的Huh7细胞10天,然后,提取存活的Huh7细胞(<5%)基因组DNA,扩增ORF对应的条形码进行高通量测序。对测序数据进行生物信息学分析后,发现已知的抗病毒蛋白IL-27和I型干扰素IFNA2,IFNA4,IFNA6,IFNA7都被明显富集,通过验证明显富集,但抗病毒功能未知的候选蛋白,作者发现TMEM120A(又名TACAN,NET29,TMPIT等)可以显著地抑制寨卡病毒的感染(图1.b)。另外,作者发现TMEM120A在U87MG(人脑星形胶质母细胞瘤)细胞中也可以显著地抑制寨卡病毒感染(图1.c,d),由于寨卡病毒对神经发育影响较大,这一结果预示着TMEM120A(大量表达或功能激活)或可以在胎儿发育早期干预寨卡病毒感染,预防小头症。
图1. 人类全基因组过表达筛选系统鉴定出TMEM120A可以抑制寨卡病毒感染; a. 全基因组过表达筛选系统流程图;b. 筛选所得候选基因的散点图;c, d. TMEM120A过表达可以抑制寨卡病毒感染U87MG细胞(c. 寨卡病毒 NS5 RNA水平;d. 寨卡病毒 NS1蛋白水平)
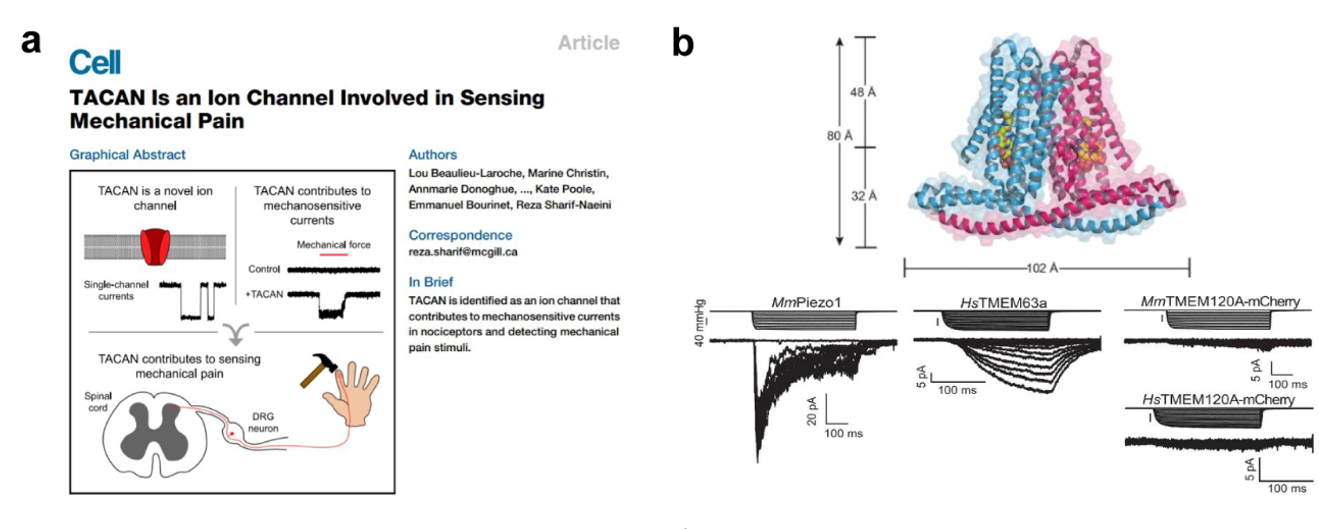
TMEM120A与2021年诺贝尔生理学或医学奖 2021年的诺贝尔生理学或医学奖被授予了David Julius和Ardem Patapoutian两位科学家,表彰他们在“发现温度和触觉受体”方面取得的突破性成就。其中,Ardem Patapoutian教授发现了可以感受机械刺激的离子通道PIEZOs(PIEZO1和PIEZO2)。有趣的是,2020年Beaulieu-Laroche, Lou等在Cell杂志报道了TMEM120A是一种全新的机械刺激感受型离子通道(图2a)【8】,有可能是继PIEZOs之后,迄今为止发现的第二类机械刺激感受型离子通道。但是,2021年陆续有来自中国(分别来自中科院生物物理所的柳振峰团队和西湖大学的吴建平团队)和美国(分别来自洛克菲勒大学的MacKinnon组和德州西南医学中心的Youxin Jiang组)4篇论文解析了TMEM120A的结构,结合生化实验均不支持TMEM120A具有离子通道活性(图2b) 【9-11】。
本研究验证了TMEM120A的疑似离子通道性质是否与其抑制寨卡病毒感染相关。作者向完全培养基中添加各种离子Ca2+,Mg2+,Mn2+,Cu2+,Zn22+,Gd3+以观察它们对寨卡病毒感染的影响,Ca2+,Gd3+的添加会整体抑制对照组和TMEM120A过表达组寨卡病毒的感染,但是并不能消除对照组和TMEM120A过表达组之间的差异。另外,2020年Cell论文中描述的离子通道的关键氨基酸的突变均不影响TMEM120A的抗病毒功能。
由于TMEM120A与PIEZOs之间可能的关联,有关TMEM120A是否是机械刺激感受型离子通道的论战或将持续,但是,
TMEM120A是通过什么机制抑制了寨卡病毒感染呢?接下来,作者进一步进行了探究。 图2. TMEM120A(TACAN)的机械刺激离子通道活性尚存在争议; a. 2020年TMEM120A首次被报道能够感受疼痛刺激的离子通道 [8];b. 2021年TMEM120A结构被报道,其机械力刺激离子通透性受到质疑 [9],Mm: Mus Musculus,Hs: Homo sapiens
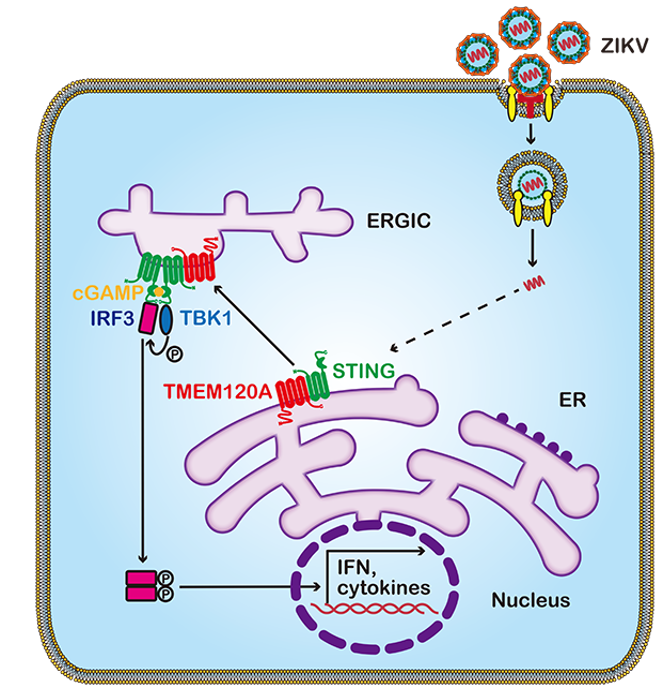
TMEM120A抑制寨卡病毒感染机制的研究之路 STING(又名TMEM173,MITA或ERIS)是细胞调动先天免疫来对抗DNA和RNA病毒不可或缺的蛋白【12】。考虑到TMEM120A与STING都位于内质网(ER)(图3a),作者探究了TMEM120A与STING的关联。本研究发现TMEM120A和STING可以共定位,且聚集呈现出大的荧光斑块(图3b),而且免疫共沉淀显示两者有直接相互作用(图3c)。进一步实验,作者发现TMEM120A过表达促进STING从ER向内质网-高尔基体中间体(ERGIC)转运(图3d),增强TBK1和IRF3的磷酸化,促进下游IFNB以及IFIT1,IFIT2,Mx1,Mx2,RSAD2,ISG15等干扰素刺激基因(ISGs)的表达,从而增强细胞整体的抗病毒先天免疫水平来抵抗病毒的感染。
值得注意的是,本研究显示TMEM120A不仅可以抑制寨卡病毒,登革病毒(DENV),黄热病病毒(YFV)等黄病毒科RNA病毒,而且可以抑制单纯疱疹病毒(HSV-1)等DNA病毒,与STING相似,具有广谱抗病毒特性。另外,作者尝试了用CRISPR-Cas9方法制备了TMEM120A敲除小鼠,发现TMEM120A全身敲除是胚胎致死的,无法用TMEM120A敲除小鼠进行病毒感染实验,所以,作者用TMEM120A敲除小鼠胚胎制备了胚胎纤维细胞(MEF),进一步验证了TMEM120A敲除会削弱细胞的先天免疫水平,促进病毒感染。
图3. TMEM120A与STING直接相互作用,促进STING转运,并激活下游信号通路; a. TMEM120A定位于ER(Calnexin为ER标记物);b. TMEM120A与STING有共定位;c. TMEM120A与STING有相互作用;d. TMEM120A促进STING从ER到ERGIC的转运
纵观整篇论文,相对于CRISPR-Cas9等功能丧失型筛选系统,作者应用人类全基因组慢病毒过表达筛选系统,鉴定出了既能抑制RNA病毒(寨卡病毒,DENV,YFV),又能抑制DNA病毒(HSV-1)感染的全新宿主蛋白TMEM120A,通过对其抗病毒机制的探究,作者发现TMEM120A可以促进STING从ER到ERGIC的转运,增强TBK1和IRF3的磷酸化,促进IFNB以及ISGs的表达,从而整体上提升宿主细胞的抗病毒先天免疫(图4)。在抗病毒功能之外,TMEM120A敲除的胚胎致死效应,提示其在胚胎发育过程中还具有其它的重要生理功能,这些谜底的揭开还有待进一步研究。
图4. TMEM120A抗病毒机制的模式图
清华大学药学院博士后李硕和钱年超为本文的并列第一作者,清华大学药学院谭旭教授为本文的通讯作者。该工作得到了科技部重点研发计划,国家自然科学基金委,北京市结构生物学高精尖中心,生物结构前沿中心和清华-北大生命科学联合中心的资助。
参考文献: 1. Dick, G., Kitchen, S. & Haddow, A. Zika virus (I). Isolations and serological specificity. Transactionsof the royal society of tropical medicine and hygiene 46, 509-520 (1952).
2. Grant, A. et al. Zika Virus Targets Human STAT2 to Inhibit Type I Interferon Signaling. Cell Host Microbe 19, 882-890, doi:10.1016/j.chom.2016.05.009 (2016).
3. Mlakar, J. et al. Zika Virus Associated with Microcephaly. N Engl J Med 374, 951-958, doi:10.1056/NEJMoa1600651 (2016).
4. Hoen, B. et al. Pregnancy Outcomes after 寨卡病毒 Infection in French Territories in the Americas. N Engl J Med 378, 985-994, doi:10.1056/NEJMoa1709481 (2018).
5. Marceau, C. D. et al. Genetic dissection of Flaviviridae host factors through genome-scale CRISPR screens. Nature 535, 159-163, doi:10.1038/nature18631 (2016).
6. Savidis, G. et al. Identification of Zika Virus and Dengue Virus Dependency Factors using Functional Genomics. Cell Rep 16, 232-246, doi:10.1016/j.celrep.2016.06.028 (2016).
7. Wang, S. et al. Integrin alphavbeta5 Internalizes Zika Virus during Neural Stem Cells Infection and Provides a Promising Target for Antiviral Therapy. Cell Rep 30, 969-983 e964, doi:10.1016/j.celrep.2019.11.020 (2020).
8. Beaulieu-Laroche, L. et al. TACAN is an ion channel involved in sensing mechanical pain. Cell 180. 5, 956-967 (2020).
9. Rong, Y. et al. TMEM120A contains a specific coenzyme A-binding site and might not mediate poking-or stretch-induced channel activities in cells. Elife 10, e71474 (2021).
10. Niu, Y. et al. Analysis of the mechanosensor channel functionality of TACAN. Elife 10, e71188 (2021).
11. Xue, J. et al. TMEM120A is a coenzyme A-binding membrane protein with structural similarities to ELOVL fatty acid elongase. Elife 10, e71220 (2021).
12. Ma, Z. & Damania, B. The cGAS-STING Defense Pathway and Its Counteraction by Viruses. Cell Host Microbe 19, 150-158, doi:10.1016/j.chom.2016.01.010 (2016).