
-

����ͨ��
����ץס�����Ƽ�
����������
����ͨ��
����ץס�����Ƽ�
����������
֬��ϸ���������д����۵��ĵ��״����»���
�����壺 �� �� С �� ʱ�䣺2023��06��06�� ��Դ���廪����ѧԺ
�༭�Ƽ���
����ϸ��ͨ�����ֲ�ͬ�ĵ����������ƻ��ƣ�Protein Quality Control Mechanisms����ֹ�к����ۼ��� ��Щ�����������ƻ��Ƶ��˻�����ϵ����ڶ����༲������������֢���������꼲���ȡ���������Ϊ�������۵����������Ƶ���Ҫ���������������������� (ERAD) ������������ ��ER-phagy�����ֻ��ƽ�����ס�����ERAD ������Եؽ����������еĵ��ף���ER-phagy����Ϊ���� (autophagy) ��һ�֣�������Ϊ������������Ƭ����...
ϸ��ͨ�����ֲ�ͬ�ĵ����������ƻ��ƣ�Protein Quality Control Mechanisms����ֹ�к����ۼ��� ��Щ�����������ƻ��Ƶ��˻�����ϵ����ڶ����༲������������֢���������꼲���ȡ���������Ϊ�������۵����������Ƶ���Ҫ���������������������� (ERAD) ������������ ��ER-phagy�����ֻ��ƽ�����ס�����ERAD ������Եؽ����������еĵ��ף���ER-phagy����Ϊ���� (autophagy) ��һ�֣�������Ϊ������������Ƭ�������������������� ��ER fragments/domains���� Ȼ������Ϊ��������Ҫ�ĵ�������ƣ� ���Ƕ�ERAD��ER-phagy�����û��ƣ������������������¶������Эͬ��ά����������̬��֪֮���١�
2023��5��31�գ� ��Ъ����ѧҽѧԺ������ڿ����鼰�廪��ѧ������ѧԺ���廪-IDG/������Կ�ѧ�о�Ժ�����������ڿ������ڡ���ȻͨѶ����Nature Communications����־�����߷�����Ϊ��֬��ϸ�������������������۵������ơ��� The mechanisms to dispose of misfolded proteins in the endoplasmic reticulum of adipocytes �����о����ġ��о���ʾ����С��֬��ϸ����SEL1L-HRD1 ERAD��ER-phagy�Ĺ�ͬ���û��ƣ�������֬��ϸ���Դ����۵��������������еĻ��ۿ��ܾ��и߶���Ӧ�ԡ�����״̬��ER-phagy�����ܵ����ƣ�SEL1L-HRD1 ERADΪ��������۵������Ļ��ơ� ��SEL1L-HRD1 ERAD�����½�ʱ��֬��ϸ����ER-phagy�������ӣ��Ӷ����ٴ����۵������γɵľۼ������������л��ۡ�����SEL1L-HRD1 ERAD ��ER-phagy ���ܾ��½�������£������۵��ĵ����Ա�������һ������������Ƭ�ۼ��ں��γɵ�����ϸ���ṹ�С�����ϸ���ṹ������ΪCoalescence of ER Fragments (CERFs)�� �о����Ʋ���ֽṹ���ܶ�֬��ϸ�� �������ڶ����ڣ���Ӧ�����������ƻ���ȱ�ݾ�����Ҫ���á�
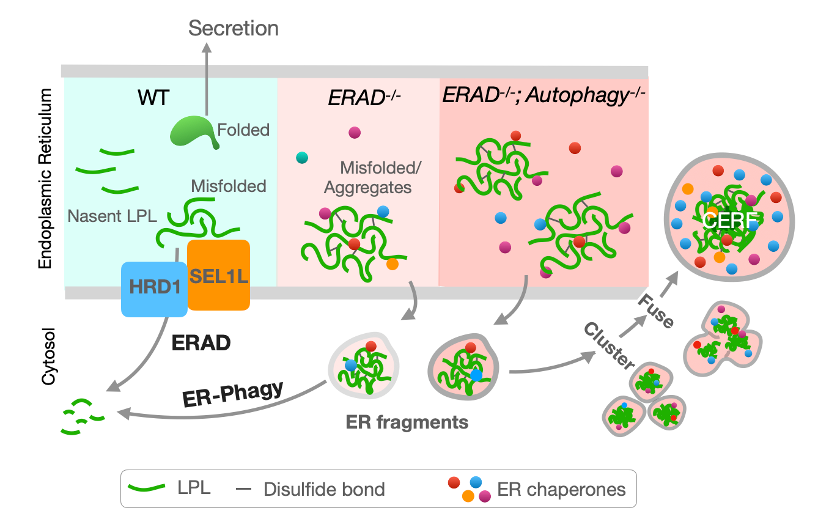
ͼ1 �C С��֬��ϸ���в���ش����������д����۵��������߾���ļ���ģ�͡�

ͼ2 �C �����������TEM���¹۲쵽��֬��ϸ������SEL1L-HRD ERAD ��ER-phagy ˫�ó�(DKO)��С���֬����֯���γɵ�CERFs�ṹ ��a�� ��ͷ����CERFs�е���Ĥ�ṹ����ʹ�� ���߽�Ⱦɫ����TEM�¹۲쵽CERFs�ṹ (��ɫ����) ������������ǻ��ǵ���BiP ��b���� LD: lipid droplet.
����о����������߳���������immunoprecipitation-mass spectrometry��������С��֬����֯��CERFs����Ҫ�ɷ֡����һϵ���������������Ƿ���֬����֬��ˮ��ø- lipoprotein lipase ��LPL����CERFs��Ҫ�ĵ��׳ɷ֣�Ҳ��֬��ϸ������Ҫ��ERAD���
LPL ��Ҫ��֬���뼡��ϸ�����ڣ�����Ҫ������ˮ������������Chylomicrons���ͼ����ܶ�֬����(VLDL)��ĸ���������LPL �Ĺ���ȱʧ�����ص�����¿��Ե���I��֬����Ѫ֢��������̶�������������֬���״�л�����ҡ���ˣ�����LPL������ϸ���ڵĺϳɻ��ƽ�Ϊ�˽⡢Ԥ�������Ƹ�Ѫ֬����������Ҫ���ס�
�о����֣�LPL���������е��۵���Ҫ��SEL1L-HRD1 ERAD���ơ�ERAD�ɼ�ʱ������������д����۵���LPL���ף��Ӷ�ȷ����ȷ�۵���LPL���ױ��������ڡ���ERAD����ȱʧ����£�LPL���γɵ��߾��Ȼ������ʱ֬��ϸ���ɼ���ER-phagy��ΪERAD ȱʧ�IJ������ƣ�����LPL���������оۼ�����֬��ϸ����ERAD��ER-phagy����˫��ȱʧ�������۵�LPL ������������CERFs, ������ʱ���γɵ��ܽ�ȵĸ߾��
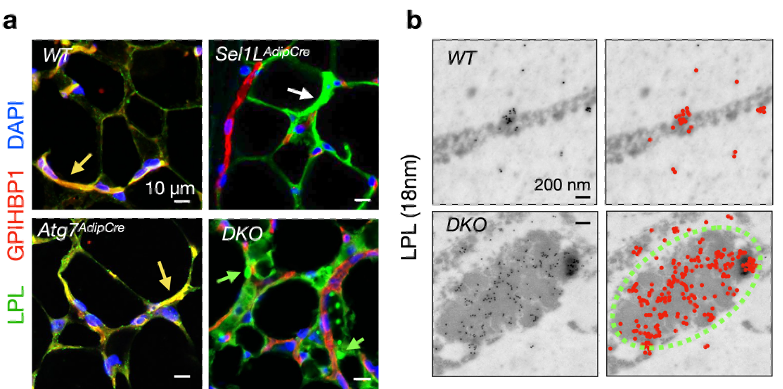
ͼ3 �C ʹ������ӫ��ʵ���ڹ��۽������¹۲쵽����Ұ����(WT)�����ɵ���(Atg7AdipCre)��С���У�LPL�ɱ�֬��ϸ�����ڲ�����Ѫ���У��Ӷ���Ѫ�ܱ�ǵ���GPIHBP1����λ��a,��ɫ��ͷ���� Ȼ����SEL1L-HRD1 ERAD����(Sel1LAdipCre)��ERAD������˫��(DKO)��С����, LPL�ۻ���֬��ϸ���дӶ����ܱ��������ڵ�Ѫ����(a, ��ɫ��ͷ)�������SEL1L-HRD1 ERAD���ã�Sel1LAdipCre)����˫��С��(DKO)�У�LPL������֬��ϸ�����ۻ��������γ�Բ�ν��㣨a, ��ɫ��ͷ����ʹ�����߽�Ⱦɫ������LPL����TEM�¹۲쵽LPL��˫��֬��ϸ���и�����CERFs�ṹ (��ɫ����)�У�b����
�о���������CERFs�е�������ֲ���̬��ӫ������߽�羵����������ʾ��CERFs�У�LPL���������еķ��Ӱ���BiP������Ϊ����֤CERFs��װ�Ļ��ƣ��о��߶�CERFs�����������ع��������ʾ�������� LPL�������Ծۼ����������ܹ��������Է�����벢�γ��пյľ������ƽ�״���ʵ�Һ�Ρ�����, ������BiP����LPLһ������벢�������γ�BiP����LPL��Һ�Ρ���Щ���ֽ�Ϊδ����һ������LPL��ϸ���ڵľۼ��͵��ػ��ƴ��»�����
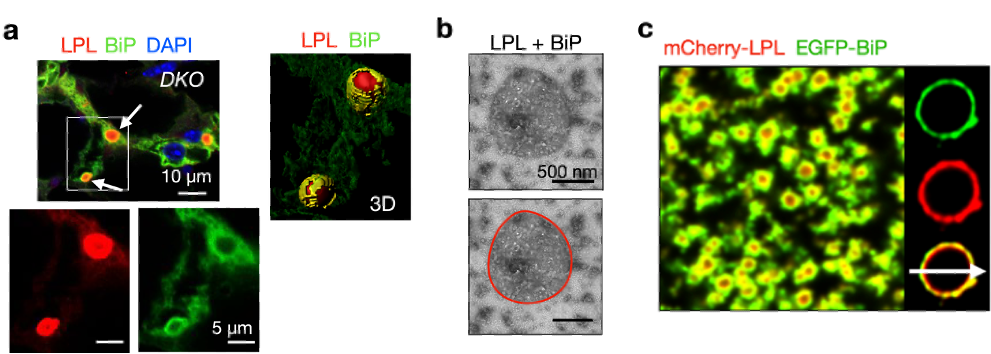
ͼ4 �C ʹ������ӫ��ʵ���ڹ��۽������¹۲쵽����ERAD������˫�õ�С��(DKO)֬����֯�У�LPL CERFs����Χ��BiP ���� ��a����ͷ������Ⱦɫ����TEM�¹۲쵽����BiP��LPLһ������루b����ӫ��Ⱦɫ���ִ�����BiP����LPLҺ�Σ�c����
���ϣ����о�����С��ģ���״�֤ʵ��SEL1L-HRD1 ERAD ��ER-phagy�����������µĻ��������߹�ͬ������Ҫ�������ܵ����۵��뽵�⣬�Ӷ�ά����������ϸ����̬�� ͬʱ���о����״η���֬��ϸ��������������Ƭ�ۼ��ں��γɵ�����ϸ���ṹCERFs, ��ʾ�����������ܣ���������CERFs����Ҫ���׳ɷּ���װ���ơ�δ���о�����һ��̽�� ERAD��ER-phagy ��CERFs�γ����ɵ����۵�ȱ������ļ����е���Ҫ�ԡ�
��Ъ����ѧҽѧԺ�������, �廪��ѧ������ѧԺ���廪-IDG/������Կ�ѧ�о�Ժ������Ϊ���Ĺ�ͬͨѶ���ߡ���Ъ����ѧ��ʿ����˪�̣�Alivia��Ϊ��һ���ߡ��廪��ѧ������ѧԺ�곿���������������鲿�֡�������Ъ����ѧҽѧԺ����ʵ���ҿ���ͬ��Э��ʵ�顣
ԭ�����ӣ�https://www.nature.com/articles/s41467-023-38690-4
����ͨ�Ź��ں�
֪����ҵ��Ƹ

