综述:F-box蛋白在癌症中的作用:从癌细胞到肿瘤微环境
《Cell Communication and Signaling》:F-box proteins in cancer: from cancer cells to the tumor microenvironment
【字体:
大
中
小
】
时间:2025年10月15日
来源:Cell Communication and Signaling 8.9
编辑推荐:
本综述系统阐述了F-box蛋白通过泛素化修饰在癌症中的多维调控作用。文章突破传统视角,不仅涵盖癌细胞自身行为(增殖、死亡、代谢重编程等),更拓展至肿瘤微环境(TME)调控,揭示了F-box蛋白在肿瘤免疫、血管生成及癌相关成纤维细胞(CAF)相互作用中的关键角色。作者强调了其作为E3泛素连接酶SCF复合物的底物识别模块,通过复杂的调控层级网络,在癌症治疗靶点开发(如PROTAC技术)中展现巨大潜力。
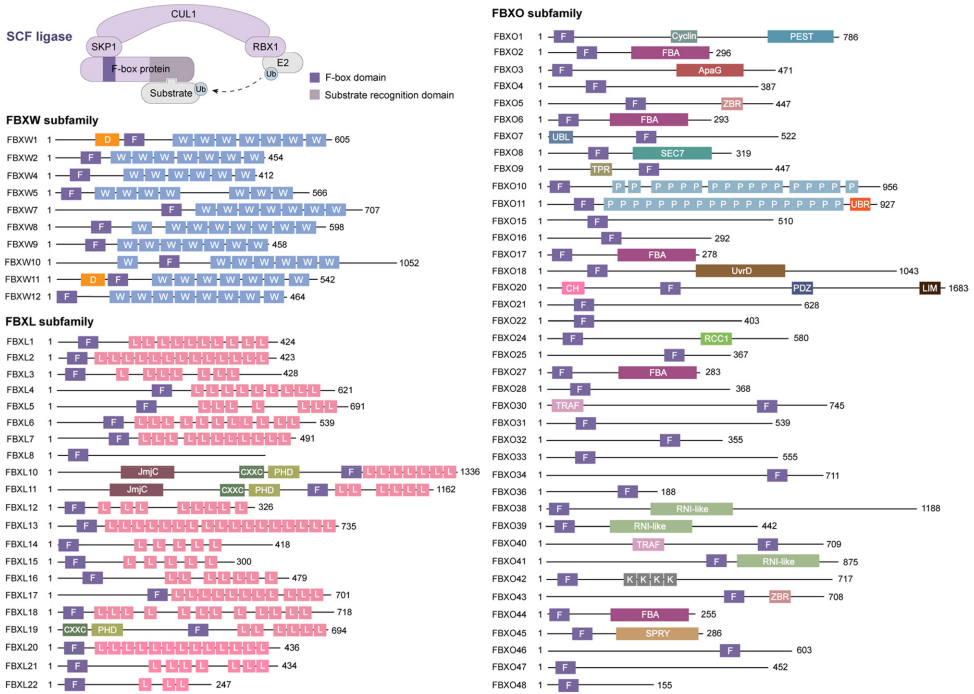
泛素化是一种关键的蛋白质翻译后修饰,通过E1、E2和E3酶的三步酶促反应,将泛素分子标记到底物蛋白上。这一过程不仅导致经典的蛋白酶体或溶酶体降解,还能通过不同连接类型(如K48、K63)的泛素链介导非降解性结局,如调控蛋白质相互作用和定位。E3连接酶,尤其是Cullin-RING连接酶(CRL)家族中的SCF(SKP1-CUL1-F-box蛋白)复合物,通过其可变组件――F-box蛋白来特异性识别底物,从而决定了泛素化的底物特异性。
人类基因组编码约70个F-box蛋白,根据其蛋白相互作用域可分为三大亚家族:FBXW(含WD40重复结构域,10个成员)、FBXL(含亮氨酸重复结构域,21个成员)和FBXO(仅含F-box结构域及其他未特征化结构域,39个成员)。它们通过识别底物蛋白上特定的降解子(degron)来发挥作用,其中磷酸化修饰是经典的识别模式,例如FBXW7识别CDC4磷酸化降解子(CPD)。此外,糖基化等修饰以及底物蛋白的四级结构也作为降解子被识别,如FBXO22和FBXL17可分别识别转录因子BACH1的不同四聚体结构降解子,实现了对同一底物的动态互补调控。
除了调控传统的癌细胞增殖、迁移、侵袭和凋亡外,F-box蛋白在癌症的新兴生物学特征中也扮演着重要角色。
癌细胞表型可塑性是肿瘤异质性和适应性的基础,主要包括上皮-间质转化(EMT)、去分化(获得肿瘤干细胞CSC特性)和转分化。F-box蛋白是这一过程的多功能开关。
在EMT过程中,FBXO22、FBXO31、FBXW1和FBXW7等可靶向降解关键转录因子SNAIL;FBXO28、FBXO45和FBXL14则分别靶向SLUG、ZEB1和TWIST1,从而抑制EMT。相反,FBXO32通过介导CtBP1的K63连接泛素化促进其核滞留,进而触发促EMT基因表达。在干细胞特性方面,FBXO15和FBXW8通过降解SOX2和NANOG抑制干细胞特性,而FBXW2则通过降解MSX2(SOX2的转录抑制因子)来增强干细胞特性。值得注意的是,FBXO8可同时抑制EMT却促进干细胞特性,提示F-box蛋白可能通过剂量依赖方式决定特定的EMT状态,从而调节CSC特征。在转分化中,FBXL10通过表观遗传学机制维持滑膜肉瘤细胞的恶性神经样表型,而FBXO7则通过稳定Rbfox2促进胶质母细胞瘤的间质转化。
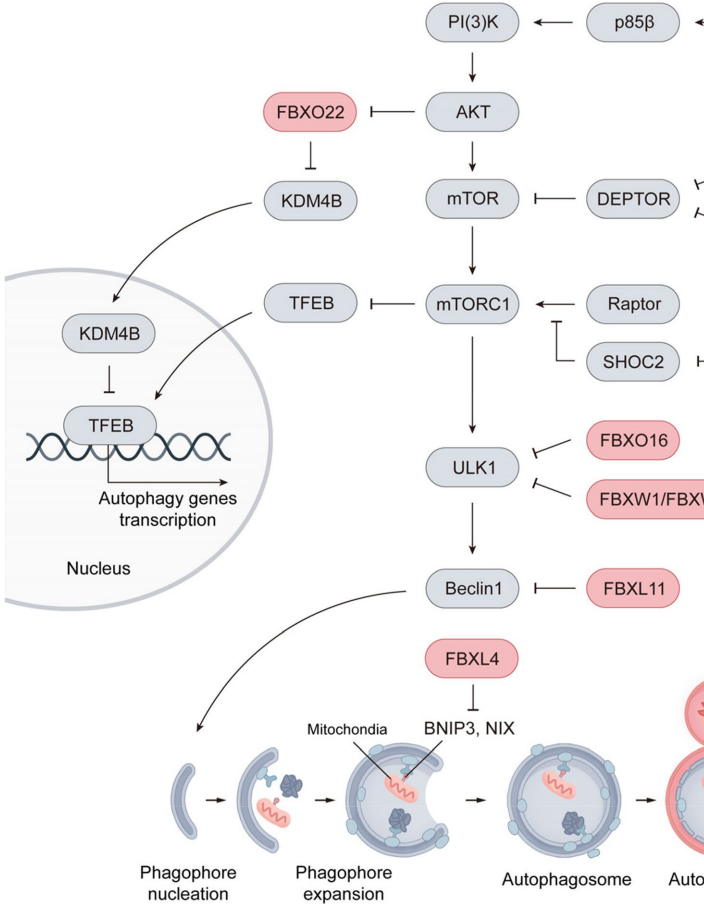
自噬在癌症中扮演双重角色。F-box蛋白通过调控自噬的不同阶段来影响癌细胞命运。在非选择性自噬中,FBXW1/FBXW11通过降解DEPTOR、FBXW7通过降解SHOC2、FBXL2通过降解p85β来激活mTORC1,从而抑制自噬。FBXO16直接降解ULK1,FBXL11降解Beclin 1,分别抑制自噬起始和吞噬泡成核。在选择性自噬如线粒体自噬(mitophagy)中,FBXW1/FBXW11通过降解ULK1,FBXL4通过降解受体BNIP3和NIX来抑制线粒体自噬。尽管FBXO22在正常细胞中通过降解KDM4B促进TFEB转录以增强自噬,但在癌细胞中,过度活化的AKT信号通路会阻断此过程。
除了凋亡,F-box蛋白也调控新兴的细胞死亡方式。在铁死亡(ferroptosis)中,FBXW7通过降解VDAC3或转录抑制NR4A1-SCD1轴促进铁死亡;FBXL2和FBXO31则分别通过降解AR和直接降解GPX4来促进铁死亡。相反,FBXO10通过降解ACSL4抑制铁死亡。在自噬性细胞死亡中,FBXW5通过降解AQP3抑制该过程,而FBXO41则通过降解FBXL1诱导自噬性细胞死亡。F-box蛋白在细胞焦亡(pyroptosis)中的作用在癌症中尚不清楚。
F-box蛋白广泛参与癌细胞的代谢重组。在糖酵解(Warburg效应)中,FBXW7通过直接降解ENO1、mTOR或c-Myc来抑制糖酵解,而FBXL1则通过K63连接泛素化激活AKT促进糖酵解。在脂质生成中,FBXW7β降解FASN,FBXO11通过降解hnRNP A2/B1间接抑制FASN、ACC1和ACLY的表达。在氨基酸代谢中,FBXO7通过降解PRMT1抑制丝氨酸合成,而FBXL10则通过表观遗传学机制增强c-Myc和ATF4的表达,促进丝氨酸-甘氨酸-一碳代谢和谷氨酸代谢。
为应对氧化应激,FBXL1通过K63连接泛素化稳定MTH1,而FBXO22和FBXL17则协同靶向不同结构降解子促进BACH1降解,激活抗氧化反应。在内质网应激中,FBXO5通过降解RNF183抑制凋亡,而ER应激通过下调FBXL1,稳定FBXW1,进而降解CDK4导致细胞周期停滞。
F-box蛋白通过影响癌细胞干性和死亡等方式调控治疗抵抗。FBXW7通过降解MCL1抑制对化疗(如顺铂)和靶向治疗(如奥希替尼)的耐药性;相反,FBXO45通过降解FBXW7促进化疗耐药。FBXL1抑制对顺铂和CHK1抑制剂的耐药性。FBXW2抑制乳腺癌对紫杉醇的耐药性,却增强对他莫昔芬的耐药性。在放疗抵抗中,FBXW7起抑制作用,而FBXO22和FBXO6则促进放疗抵抗。
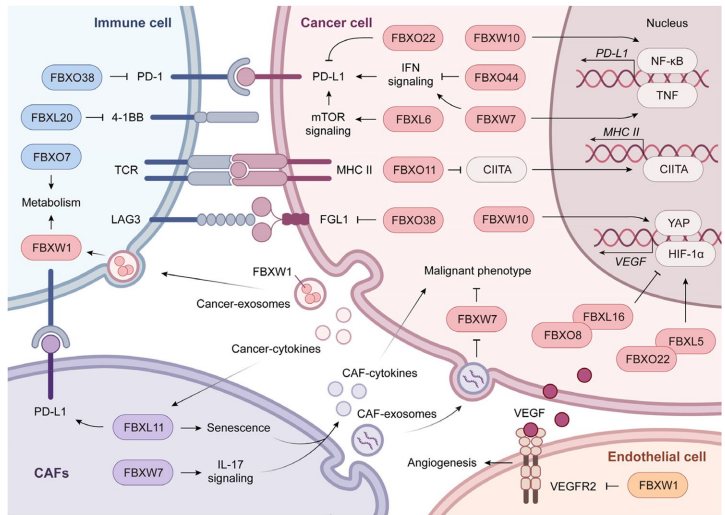
F-box蛋白通过多种机制重塑TIME并影响免疫治疗疗效。在癌细胞中,FBXW7通过维持病毒感应和干扰素信号促进PD-L1表达,增强抗PD-1疗法敏感性;而FBXO44则起相反作用。FBXO22直接降解PD-L1。FBXO11通过降解CIITA抑制MHC II类分子表达,削弱抗原呈递。在免疫细胞中,FBXO38通过降解PD-1维持T细胞抗肿瘤活性,并通过激活IL15R信号增强NK细胞功能。相反,FBXL20通过降解共刺激受体4-1BB削弱T细胞免疫力。此外,癌细胞来源的外泌体可将FBXW1递送至CD8+ T细胞,降解YAP1导致T细胞耗竭。
F-box蛋白在VEGF信号通路的不同阶段调控血管生成。FBXW7、FBXL16和FBXO8可直接降解HIF-1α;FBXL5通过降解CITED2释放其对CBP/p300的占据,增强HIF-1α转录活性;FBXO22通过降解RPS5激活PI3K/AKT信号上调HIF-1α和VEGF。FBXW10通过降解LATS2激活YAP促进VEGF产生。在终端阶段,FBXW1可降解内皮细胞上的VEGFR2抑制血管生成。
F-box蛋白与癌相关成纤维细胞(CAF)-癌细胞相互作用
F-box蛋白是CAF与癌细胞双向通讯的核心协调者。缺氧可下调CAF中的FBXW7,增强KMT2甲基转移酶活性,增加H3K4甲基化和IL-6、CXCLs等细胞因子的分泌,从而促进癌细胞迁移和侵袭。癌细胞来源的IL-6和TNF-α可上调正常成纤维细胞中的FBXL11,使其转化为CAF,并诱导衰老相关分泌表型(SASP),释放促癌因子。CAF来源的外泌体可抑制癌细胞中FBXW7的表达,稳定其底物如β-连环蛋白(β-catenin),增强干细胞特性、EMT、转移和耐药性。细胞外基质(ECM)硬度可减弱FBXW1/FBXW11与YAP的相互作用,促进癌细胞增殖和EMT。
F-box蛋白通过其庞大的底物库,在癌细胞和TME中构建了一个广泛且多层次的调控网络,精细调控着癌症的各个方面。其功能具有上下文依赖性,并受到不同层面的调控。虽然部分F-box蛋白的功能已被深入探索,但许多成员的作用仍不清楚。鉴于其功能多样性和关键作用,F-box蛋白已成为癌症治疗(如PROTAC和分子胶技术)极具潜力的靶点。然而,由于其底物多样性可能带来脱靶效应,未来开发F-box蛋白靶向疗法仍需谨慎,并需要更多功能和结构证据的支持。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号