
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
综述:感知还原力以决定细胞命运:信号转导中的黄素氧化还原开关
《Current Microbiology》:Sensing the Reducing Power to Determine the Cell Fate: Flavin Redox-Switches in Signal Transduction
【字体: 大 中 小 】 时间:2025年10月22日 来源:Current Microbiology 2.6
编辑推荐:
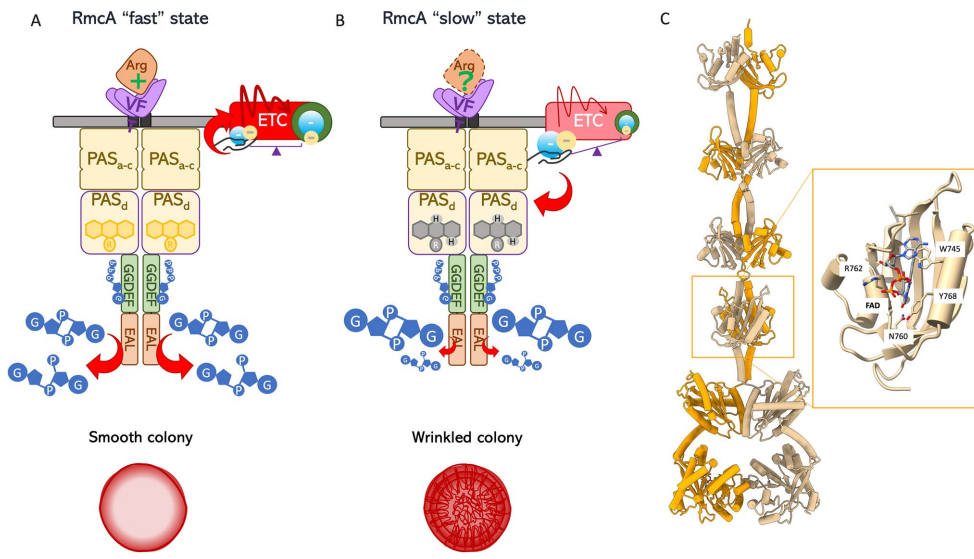
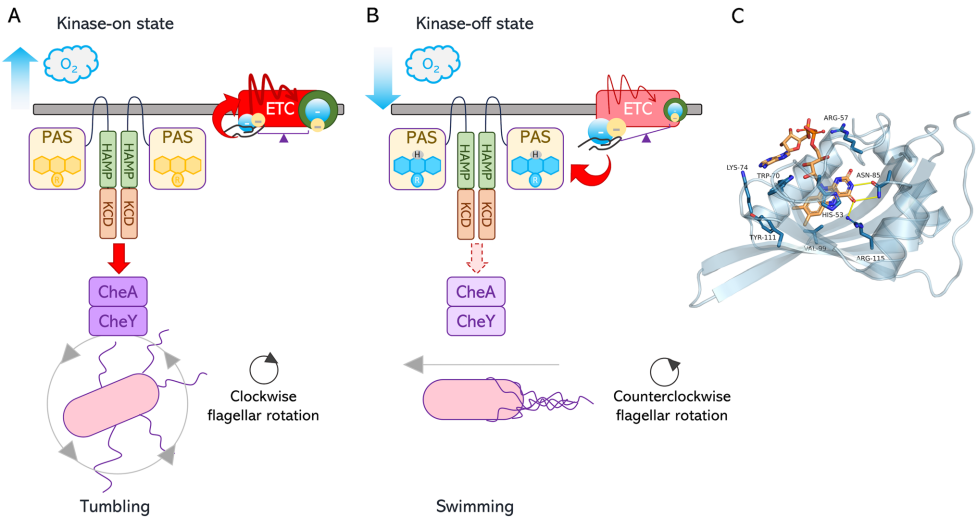
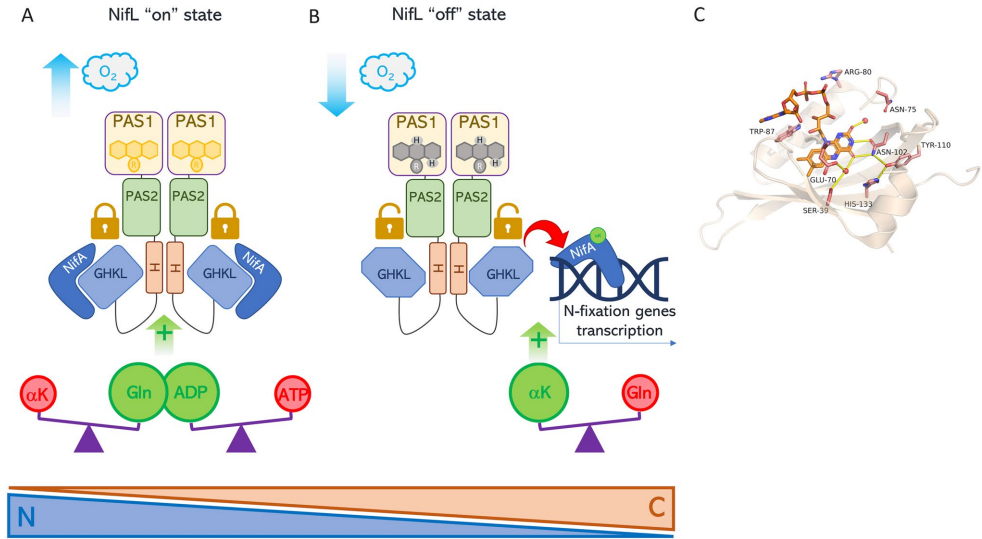
本综述深入探讨了黄素辅因子(特别是FAD/FADH2)作为细菌细胞内关键氧化还原传感器,通过感知还原力(Reducing Power)变化来调控细胞命运的核心机制。文章系统阐述了四种代表性黄素依赖的单组分信号转导系统(RmcA, AxDGC2, Aer, NifL)如何整合环境信号(如氧气、精氨酸)与胞内代谢状态(如c-di-GMP水平、ATP/ADP比值、C/N比),进而精细调控生物膜形态、能量趋化性及固氮作用等关键生物学过程,为开发针对细菌感染(如铜绿假单胞菌)的新型电疗(Electroceutical)策略提供了分子理论基础。
 生物通微信公众号
生物通微信公众号

