通过双亲和性蛋白Pitchoune介导的核仁与异染色质凝聚体层级相互作用调控细胞核高级结构
《Nature Cell Biology》:Hierarchical interactions between nucleolar and heterochromatin condensates are mediated by a dual-affinity protein
【字体:
大
中
小
】
时间:2025年11月25日
来源:Nature Cell Biology 19.1
编辑推荐:
本研究针对真核细胞核内两大无膜细胞器―核仁与着丝粒周围异染色质(PCH)的空间共组织机制这一前沿科学问题,通过果蝇胚胎发育高分辨率活体成像、分子动力学模拟与基因操作,首次揭示了DEAD-box RNA解旋酶Pitchoune(Pit)作为双亲和性分子 linker,通过其PxVxL基序与HP1a二聚体相互作用,介导了PCH-核仁层级组装的动态过程。该研究阐明了生物大分子凝聚体之间通过亲和性层级和双亲和性分子进行高级组织的物理原理,对理解核架构在发育、衰老与疾病中的作用具有重要意义。
在真核细胞的细胞核内,存在着许多没有膜结构的功能性区室,它们被称为生物分子凝聚体(biomolecular condensates),其形成往往与液-液相分离(liquid-liquid phase separation, LLPS)机制密切相关。其中,核仁(nucleolus)和着丝粒周围异染色质(pericentromeric heterochromatin, PCH)是细胞核中体积最大的两种凝聚体。核仁是核糖体合成的工厂,而PCH对于染色体分离、基因组稳定性和转录沉默至关重要。在显微镜下,我们常常观察到PCH紧密地环绕在核仁周围,这种保守的空间关联暗示着两者之间存在着精密的调控机制,然而,这种高级核结构是如何建立并维持的,其背后的分子原理至今仍不清晰。
为了回答这一基本细胞生物学问题,一项发表在《Nature Cell Biology》杂志上的研究,利用果蝇(Drosophila melanogaster)胚胎发育这一理想模型,结合高分辨率活体成像、遗传操作、生物化学分析和计算模拟等多种技术,深入探究了核仁与PCH动态互作的机制。研究人员发现,一种名为Pitchoune(Pit)的DEAD-box RNA解旋酶扮演了关键角色,它如同一个“分子链接器”,通过其不同的结构域分别与核仁组分和PCH结合,从而介导了这两种巨大凝聚体的层级式组织。
研究的关键技术方法包括:利用果蝇胚胎进行高分辨率活细胞时间推移成像,动态追踪PCH(标记蛋白HP1a)和核仁各亚区室(标记蛋白Fibrillarin, Fib,代表致密纤维组分Dense Fibrillar Component, DFC;Modulo, Mod,代表颗粒组分Granular Component, GC)在早期发育过程中的空间组织动态;通过遗传手段剔除核糖体DNA(rDNA)以消除功能性核仁,观察PCH的后续变化;利用荧光原位杂交(FISH)技术定位特定DNA重复序列;结合基于多相润湿理论(multiphase wetting theory)的粗粒度分子动力学模拟(coarse-grained molecular dynamics simulations),构建理论模型预测其组织原理;并通过RNA干扰(RNAi)、免疫共沉淀(Co-Immunoprecipitation, Co-IP)、蛋白质纯化与体外结合实验等分子细胞生物学方法,验证了Pit与PCH标记蛋白HP1a之间的直接相互作用,并确定了其相互作用的关键基序。
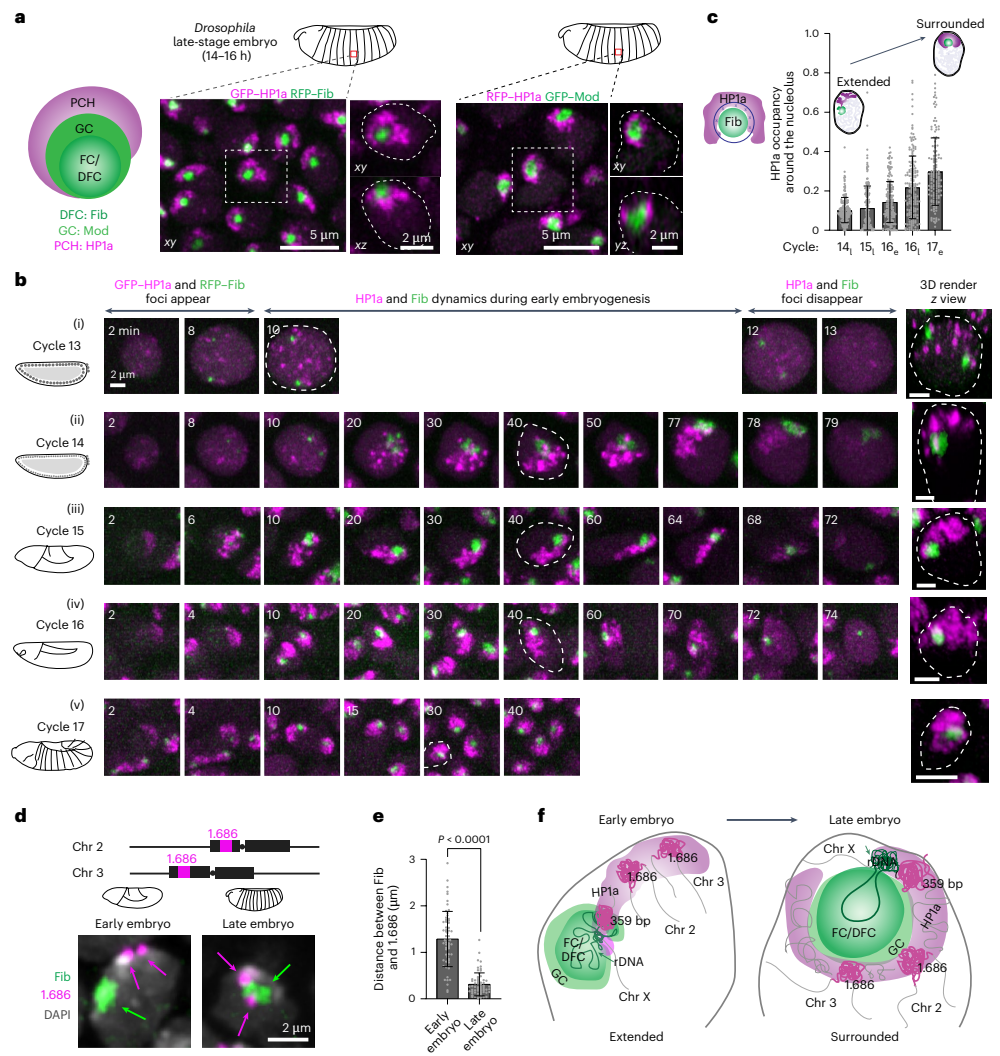
动态de novo组装果蝇胚胎发育过程中核仁周围的PCH组织
研究人员首先通过活体成像精确描绘了果蝇胚胎发育晚期PCH与多相核仁的三维组织。他们发现,PCH主要组织在核仁的GC区域周围,形成一种典型的“环绕”构象。然而,这种结构并非一成不变。通过追踪早期胚胎核 cycles 13至17的动态过程,他们观察到PCH的组织经历了从沿核边缘延伸的“延伸构象”,到通过rDNA一端锚定在核仁上,最终动态转变为稳定包裹核仁的“环绕构象”的过程。DNA FISH实验进一步证实,即便不携带rDNA的染色体,其PCH在发育后期也会被招募到核仁边缘。这些结果揭示了PCH-核仁关联的建立是一个高度动态、逐步发展的过程。
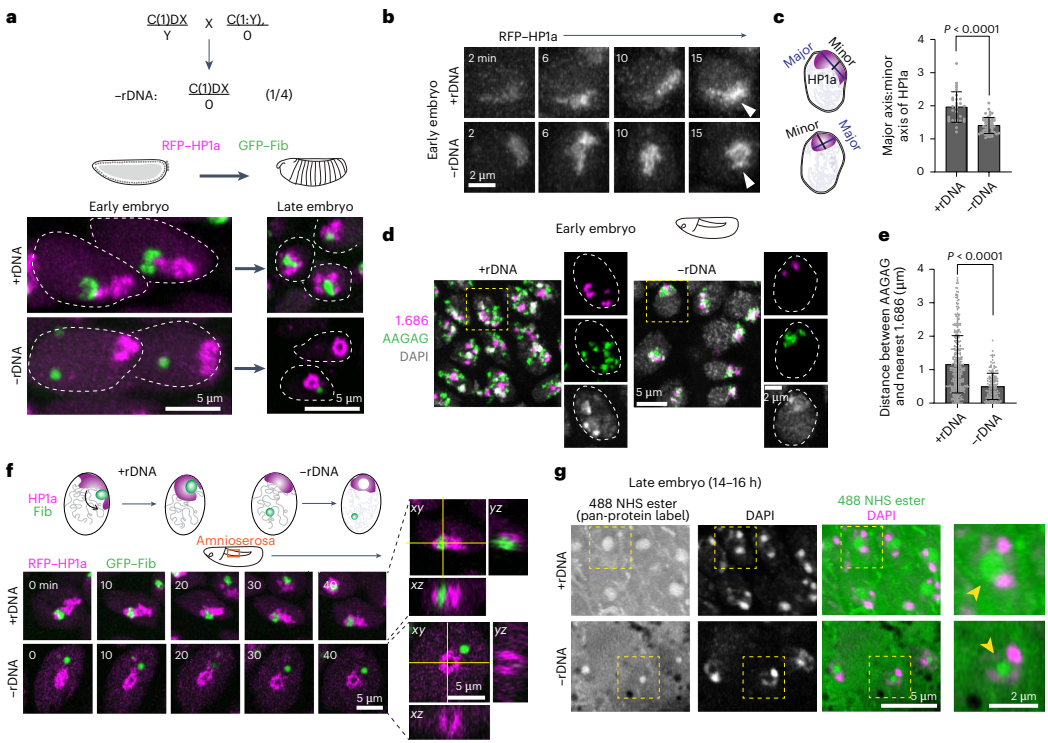
移除rDNA导致PCH压缩加剧,随后重组为环状结构并重新分布核仁蛋白
为了探究核仁对PCH组装的影响,研究人员观察了缺乏rDNA(-rDNA)的胚胎。结果发现,在-rDNA胚胎中,无法形成正常核仁,核仁蛋白Fib形成了新的球形凝聚体(neocondensates)。更为惊奇的是,PCH的表现也发生了剧变:在早期胚胎中,PCH变得更加紧凑;而在发育后期,PCH则重组为一个环状(炸面圈状)结构,其中心出现了一个缺乏HP1a的“PCH空洞”。这个空洞内不含有DNA、组蛋白修饰H3K9me2或RNA,但却富含蛋白质,提示其中可能形成了由某种(些)具有自组装能力的蛋白质构成的新凝聚体。
模拟核仁和PCH之间的层级相互作用再现了其体内组织,并提示双亲和性蛋白参与其关联
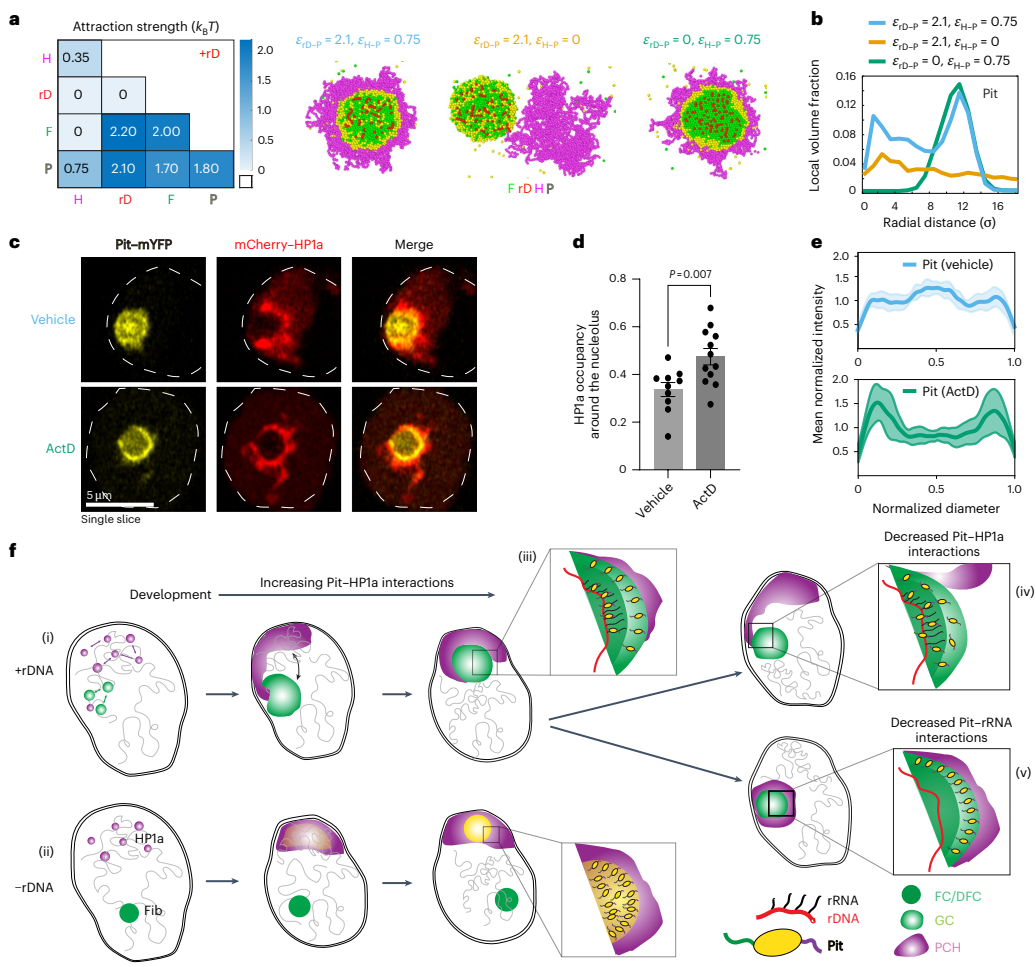
基于相分离和润湿理论,研究人员构建了一个最小化的粗粒度模型,包含PCH、rDNA、Fib和一个假设的双亲和性蛋白X。通过设定四种组分之间相互作用强度的层级关系,模拟成功复现了野生型(+rDNA)核中PCH环绕核仁的“环绕构象”,以及-rDNA核中PCH形成环状结构且内部出现蛋白X富集相的现象。模型预测,存在一种与PCH和核仁组分均有亲和力的“双亲和性”蛋白,对于维持PCH-核仁的正确空间组织至关重要。
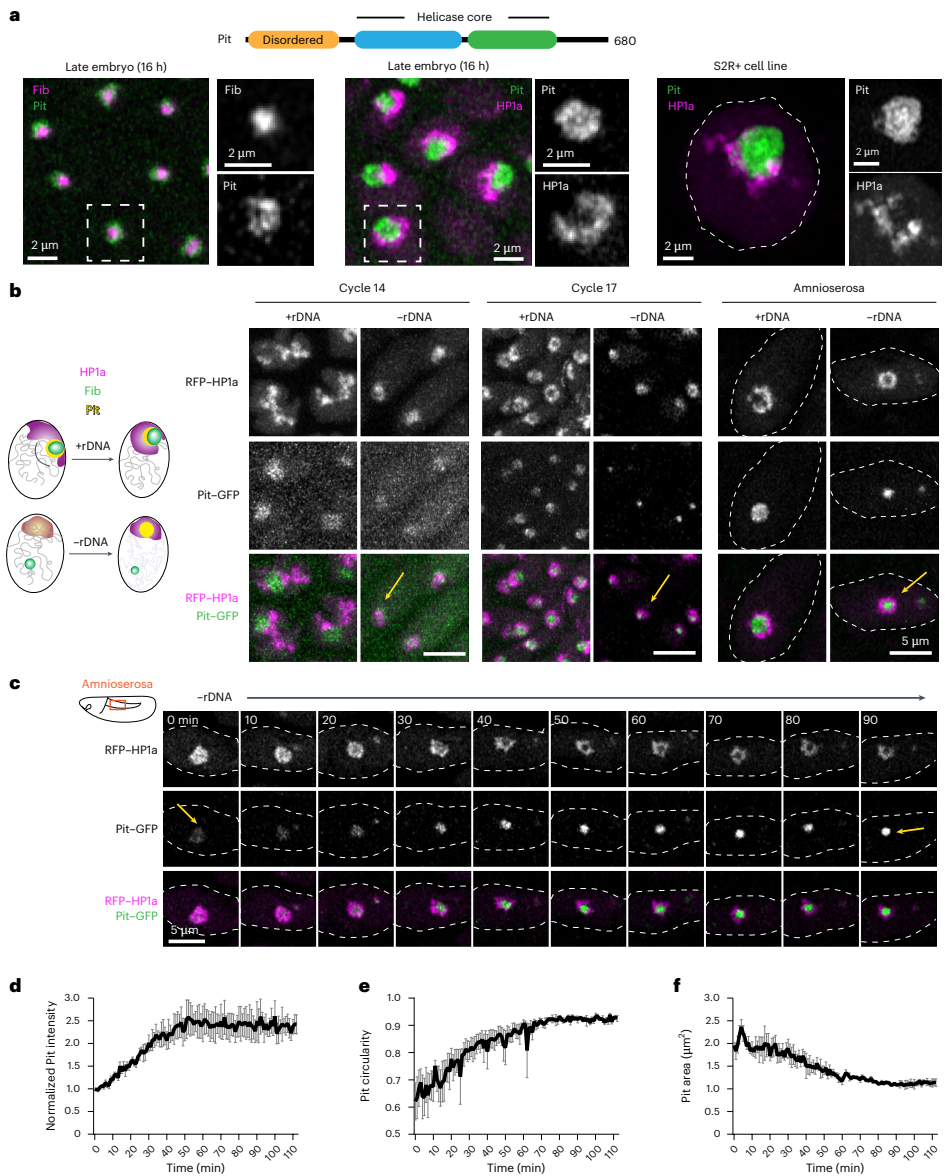
DEAD-box RNA解旋酶Pit是一种在-rDNA胚胎的PCH内形成新凝聚体的GC蛋白
那么,这个假说的双亲和性蛋白X在现实中是谁呢?前期研究提示,DEAD-box RNA解旋酶Pitchoune(Pit)在-rDNA胚胎中能形成温度依赖性的凝聚域,且其位置与紧凑的PCH域相似。本研究发现,Pit定位于核仁的GC区域。在-rDNA胚胎中,Pit的分布发生了显著变化:它最初与HP1a混合,随后逐渐分离,并在PCH环状结构的空洞内形成一个高度富集、近乎球形的蛋白质新凝聚体。这一现象与模型预测的蛋白X的行为高度吻合。
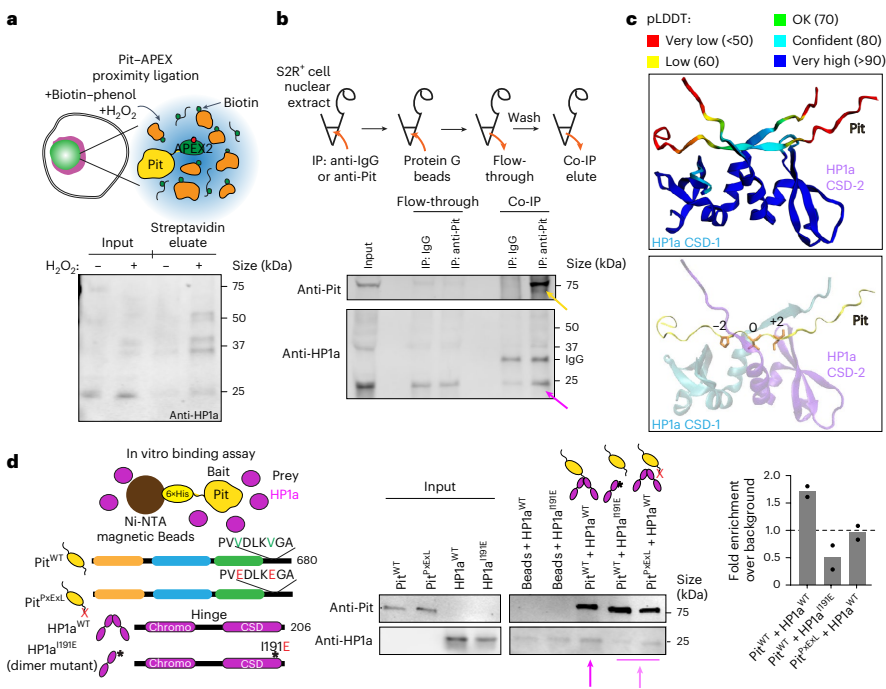
接下来的实验证实了Pit与HP1a之间存在直接物理相互作用。通过邻近标记(APEX2)、免疫共沉淀和体外Pull-down实验,研究人员发现Pit能够与HP1a结合。生物信息学分析在Pit的C端无序区鉴定出两个串联的PxVxL基序(这是已知的HP1a结合基序),AlphaFold-Multimer结构预测显示该基序可结合在HP1a二聚体的色阴影结构域(CSD)界面的疏水口袋中。体外结合实验进一步证实,Pit通过其PxVxL基序与HP1a二聚体直接相互作用,破坏该基序(PitPxExL)或HP1a的二聚化(HP1aI19E)均会显著削弱两者的结合。
Pit及其与HP1a的相互作用是PCH-核仁关联所必需的
功能实验表明,在果蝇幼虫组织或S2R+细胞中敲低Pit,会导致PCH在核仁边缘的占有率显著下降,即“环绕构象”被破坏。而回补野生型Pit可以挽救这一表型,但回补不能结合HP1a的PitPxExL突变体则无法挽救。有趣的是,回补一个解旋酶活性失活但HP1a结合能力正常的PitDQVD突变体,仍能部分恢复PCH-核仁关联。这些结果证明Pit对于建立PCH-核仁的空间关联是必需的,并且这一功能依赖于其通过PxVxL基序与HP1a的相互作用。
调节Pit与核仁或HP1a的相互作用会改变其核仁内定位和PCH-核仁关联
研究人员还发现,改变Pit的相互作用会影响其自身的核仁内定位。野生型Pit在核仁内部(FC/DFC)和外围(GC)均有富集。不能结合HP1a的PitPxExL突变体在核仁内分布均匀,失去了外围富集;而解旋酶失活的PitDQVD突变体则主要富集于核仁外围。通过精炼模型引入Pit与rDNA(代表其解旋酶活性相关的相互作用)的相互作用参数后,模拟成功复现了这些定位模式。使用低剂量放线菌素D(ActD)抑制rRNA转录,模拟破坏Pit-rDNA相互作用,同样观察到了Pit向核仁外围的重新分布以及PCH环绕程度的增加。这些结果表明,Pit在核仁内的定位及其介导PCH-核仁关联的功能,取决于其不同结构域与核仁组分(主要通过解旋酶活性)和PCH(通过PxVxL-HP1a相互作用)之间相互作用的平衡。
综上所述,这项研究揭示了细胞核内两大凝聚体―核仁与PCH―通过一种双亲和性分子链接器Pit进行层级组装的动态过程和分子机制。Pit通过其不同的结构域介导了与核仁和PCH组分强弱不同的相互作用,这种亲和性层级决定了凝聚体的最终空间构象。该研究不仅阐明了核仁-PCH这一保守核结构建立的机制,更重要的是,它提出了一个普适性的原理:不同生物分子凝聚体之间可以通过“双亲和性”分子及其相互作用强度的层级关系,实现高级的空间共组织。这一原理可能广泛适用于其他无膜细胞器的组装与互作。此外,研究还提示,在发育、细胞应激、衰老或疾病状态下,关键相互作用网络的扰动可能导致生物分子错误定位和异常凝聚体的形成,从而影响细胞功能。因此,这项工作为理解核架构在生理和病理条件下的调控机制提供了新的框架和视角。
 生物通微信公众号
生物通微信公众号
 生物通新浪微博
生物通新浪微博
今日动态 |
人才市场 |
新技术专栏 |
中国科学人 |
云展台 |
BioHot |
云讲堂直播 |
会展中心 |
特价专栏 |
技术快讯 |
免费试用
版权所有 生物通
Copyright© eBiotrade.com, All Rights Reserved
联系信箱:
粤ICP备09063491号