
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
组蛋白动态调控工具包驱动无脊椎动物基因组与发育进化的多组学解析
【字体: 大 中 小 】 时间:2025年06月11日 来源:Genome Biology 10.1
编辑推荐:
本研究通过多组学方法系统解析了环节动物组蛋白调控系统的进化特征,发现组蛋白变体H2A.X的平行进化现象及组蛋白修饰酶(HME)表达异时性与发育模式的关联,揭示了组蛋白调控工具包在动物形态多样性形成中的重要作用,为理解表观遗传调控的进化机制提供了新视角。
在生命演化的长河中,组蛋白作为真核生物基因组包装的核心元件,其动态修饰和变体替换构成了复杂的表观遗传调控网络。然而当前对组蛋白调控机制的认识主要局限于少数模式生物,这严重限制了对表观遗传变异如何驱动动物形态多样化的理解。环节动物作为螺旋动物中形态最丰富的类群之一,其发育过程中表现出的显著异时性现象――如Owenia fusiformis的摄食型幼虫与Capitella teleta非摄食型幼虫的发育时序差异,为研究表观遗传调控与发育进化提供了理想模型。
英国伦敦大学玛丽女王学院的研究团队联合剑桥Altos Labs等机构,通过对三种发育模式迥异的环节动物(O. fusiformis、C. teleta和微型基因组物种Dimorphilus gyrociliatus)开展多组学分析,系统揭示了组蛋白调控工具包的进化规律及其与发育程序的关联。这项发表在《Genome Biology》的研究首次绘制了螺旋动物组蛋白修饰的全景图谱,发现组蛋白变体H2A.X在真核生物中广泛存在的平行进化现象,并阐明了组蛋白修饰酶表达时序与生命史策略的关联。
研究主要采用四大技术方法:(1)基因组重注释结合系统发育分析鉴定组蛋白基因家族;(2)发育时序RNA-seq和ATAC-seq解析基因表达与染色质动态;(3)定量质谱技术检测组蛋白翻译后修饰(hPTMs);(4)比较基因组学分析组蛋白修饰酶(HME)的进化。样本来源于实验室培养的三种环节动物不同发育阶段个体。
【组蛋白库显示保守与特化并存】
通过最大似然法和贝叶斯系统发育重建,研究发现三种环节动物保留了双侧对称动物祖先的全套核心组蛋白(caH2A、caH2B、caH3、caH4)和变体(H2A.X、H2A.Z、macroH2A、H3.3、cenH3)。特别值得注意的是,基因组极度紧凑的D. gyrociliatus每个典型组蛋白仅保留2个拷贝,创下后生动物组蛋白最低拷贝数记录。组蛋白基因呈串联簇状排列,但不同物种的基因簇组织结构存在显著差异,如O. fusiformis染色体5上的大型基因簇包含35个组蛋白基因,呈现H4-H3-H2A-H2B的重复单元模式。

【动态开放的染色质环境】
ATAC-seq分析揭示组蛋白基因位于超开放染色质区域,在O. fusiformis和C. teleta中,原肠胚形成后染色质可及性显著增加。这种动态调控模式在D. gyrociliatus中表现不同,其开放区域仅局限于转录起始位点附近。表达分析显示典型组蛋白在早期胚胎高表达,而变体如h2az和h3-3在细胞命运决定阶段表达上调,macroh2a则在成体达到峰值,与已知的干细胞维持和终末分化功能相符。
【H2A.X变体的平行进化】
研究首次在环节动物O. fusiformis中发现两种H2A.X变体:含可磷酸化酪氨酸142的H2A.X-Y和含苯丙氨酸142的H2A.X-F。通过PHI-BLAST检索1656条真核生物H2A.X序列发现,这两种变体在脊椎动物、刺胞动物和软体动物等多个门类独立出现,其中H2A.X-F在植物中占主导地位。值得注意的是,H2A.X-F在快速分裂的早期胚胎中优势表达,暗示其可能适应快速细胞周期需求。

【组蛋白修饰酶的异时性表达】
比较基因组学分析发现,尽管D. gyrociliatus组蛋白库极度精简,但其组蛋白修饰酶(HME)基本完整且部分扩张(如PRMT6)。软聚类分析显示,约44.6%的HME在O. fusiformis和C. teleta间存在表达时序差异:如H3K27乙酰转移酶kat3a/b(crebbp/ep300)在摄食型幼虫中早期表达,而在非摄食型幼虫中延迟表达。成体质谱分析进一步证实,这些差异与H3K27ac、H3K27me3等关键hPTMs的丰度变化相关。
【PRMT6的功能创新】
在D. gyrociliatus中发现的PRMT6基因扩张尤为特殊,其中PRMT6-A和PRMT6-C通过结构域融合(如半乳糖结合凝集素结构域)产生新功能。AlphaFold3结构预测显示,这些变异体丢失了关键的酪氨酸二元体和SAM结合位点,可能演变为催化失活的调控蛋白。这种在基因组精简背景下的酶家族创新,为理解表观遗传调控的可塑性提供了新案例。
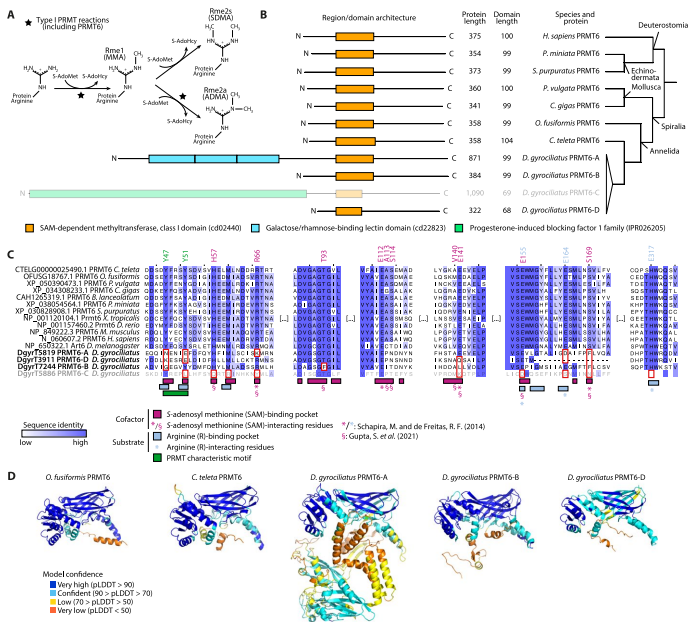
这项研究通过建立环节动物表观遗传调控的进化框架,揭示了几个关键科学发现:首先,组蛋白变体H2A.X的平行进化现象表明,表观遗传工具包的创新可能通过相似分子机制在不同谱系中反复出现;其次,组蛋白修饰酶的异时性表达与发育策略转变存在关联,为理解形态多样性产生的表观遗传基础提供了分子证据;最后,极端基因组精简背景下组蛋白系统的适应性演化(如D. gyrociliatus中PRMT6的结构创新),展现了表观遗传调控网络的惊人可塑性。
该研究的科学意义在于:技术上,建立了适用于非模式动物的多组学分析流程;理论上,提出了"组蛋白调控工具包的可塑性驱动发育进化"的新假说;应用上,为后续通过CUT&Tag等新技术在单核糖体水平解析hPTMs奠定了基础。Francisco M. Martín-Zamora等学者的工作不仅填补了螺旋动物表观遗传研究的空白,更为理解真核生物基因组调控的进化规律提供了重要范式。
生物通微信公众号
知名企业招聘

