
-

ЙъОпНЁ№ЩОў
ЕгДгЧҘЧЎЙъГьҝЖјј
Мш¶ҜөДВцІ«
ЙъОпНЁ№ЩОў
ЕгДгЧҘЧЎЙъГьҝЖјј
Мш¶ҜөДВцІ«
№ыУ¬іб°тРОМ¬ұдТмөДҝзіЯ¶ИұЈКШРФҪТКҫОўҪш»ҜУләкҪш»ҜјдөДұзЦӨ№ШПө
ЎҫЧЦМеЈә ҙу ЦР РЎ Ўҝ КұјдЈә2025Дк07ФВ08ИХ АҙФҙЈәCommunications Biology 5.2
ұајӯНЖјцЈә
ЎЎЎЎұҫСРҫҝНЁ№э·ЦОц№ыУ¬іб°тРОМ¬ФЪБщёцЙъОпЧйЦҜІгҙОЙПөДұдТмДЈКҪЈ¬ҪТКҫБЛОўҪш»ҜұдТмУләкҪш»Ҝ·Ц»ҜЦ®јдөДХэПа№Ш№ШПөЎЈСРҫҝИЛФұ·ўПЦ·ўУэФлТфЈЁFЈ©Ул4000НтДкОпЦЦ·Ц»ҜЈЁRЈ©өДПа№ШРФЧоЗҝЈ¬МфХҪБЛҙ«НіТЕҙ«ұдТмЈЁGЈ©ЦчөјҪш»ҜөД№ЫөгЈ¬Мбіц·ўУэПөНіУліӨЖЪККУҰРФІЁ¶ҜАъК·өДТ»ЦВРФЈЁcongruence hypothesisЈ©КЗҪвКНҪш»ҜҝЙФӨІвРФгЈВЫөД№ШјьЎЈёГСРҫҝОӘАнҪвІ»Н¬КұјдіЯ¶ИҪш»Ҝ№эіМөД№ШБӘМṩБЛРВКУҪЗЎЈ
ФЪҪш»ҜЙъОпС§БмУтЈ¬ОўҪш»ҜЈЁmicroevolutionЈ©УләкҪш»ҜЈЁmacroevolutionЈ©өД№ШПөТ»ЦұКЗәЛРДХщТйЎЈЧФПЦҙъЧЫәПҪш»ҜВЫТФАҙЈ¬ҙ«Ні№ЫөгИПОӘЦЦИәЛ®ЖҪөДН»ұдЎўСЎФсЎўТЕҙ«ЖҜұдөИОўҪш»Ҝ№эіМЧгТФҪвКНіӨЖЪҪш»ҜДЈКҪЎЈИ»¶шХвТ»"НвНЖ№Ыөг"ГжБЩёщұҫРФМфХҪЈәОӘәО»щУЪөұҙъТЕҙ«ұдТмЈЁGЈ©өДФӨІвіЈУліӨЖЪҪш»Ҝ№мјЈІ»·ыЈҝХвёцұ»іЖОӘ"ҝЙФӨІвРФгЈВЫ"өДПЦПуЈ¬°өКҫЧЕҪш»Ҝ№эіМЦРҝЙДЬҙжФЪЙРОҙИПЦӘөДЙоІг»ъЦЖЎЈ
ОӘҪвҝӘХвёцГХНЕЈ¬Keita SaitoөИСРҫҝХЯҝӘХ№БЛТ»ПоҝзФҪ40ёц°ЩНтДкКұјдіЯ¶ИөДҙҙРВСРҫҝЎЈНЁ№эПөНі·ЦОц№ыУ¬іб°тРОМ¬ФЪБщёцЙъОпЧйЦҜІгҙОөДұдТмДЈКҪЎӘЎӘҙУёцМе·ўУэФлТфөҪЦЦјдәкҪш»Ҝ·Ц»ҜЈ¬СРҫҝНЕ¶У·ўПЦІ»Н¬ІгҙОөДұдТмДЈКҪіКПЦҫӘИЛөДТ»ЦВРФЎЈЧоТэИЛЧўДҝөД·ўПЦКЗЈәҙ«НіИПОӘПаҫаЧоФ¶өД·ўУэФлТфЈЁfluctuating asymmetry, FЈ©УләкҪш»Ҝ·Ц»ҜЈЁRЈ©ҫ№ПФКҫіцЧоЗҝПа№ШРФЈЁR2=0.85Ј©Ј¬¶шТЕҙ«ұдТмЈЁGЈ©УлЦЦИә·Ц»ҜЈЁDpopЈ©өД№ШПө·ҙ¶шПа¶ФҪПИхЈЁR2=0.58Ј©ЎЈХвР©·ўПЦЦ§іЦБЛ"Т»ЦВРФјЩЛө"ЈЁcongruence hypothesisЈ©Ј¬јҙ·ўУэПөНіІъЙъұдТмөДДЬБҰ·ҙУіБЛіӨЖЪККУҰРФ·еЦөөДІЁ¶ҜАъК·Ј¬¶шІ»ҪцКЗөұҙъТЕҙ«ФјКшЎЈ
СРҫҝІЙУГ¶аО¬¶ИјјКх·Ҫ·ЁЈә1) ІЙјҜ14ёц№ыУ¬ОпЦЦЈЁ°ьАЁD. simulansәНD. lutescensЈ©өДіб°тСщұҫҪЁБўөИҙЖПөЈЁisofemale linesЈ©Ј»2) НЁ№эјёәОРОМ¬ІвБҝ·ЁЈЁgeometric morphometricsЈ©Бҝ»Ҝ12ёцібВцұкЦҫөгөДЧшұкұдТмЈ»3) К№УГMCMCglmmұҙТ¶Л№»мәПДЈРН№АјЖБщёцІгҙОөДРӯ·ҪІоҫШХуЈЁR, Dsp, Dpop, G, E, FЈ©Ј»4) Ҫ«Н»ұд·ҪІоҫШХуЈЁMЈ©Ул·ўУэФлТфҫШХуЈЁFЈ©ҪшРР»Ш№йұИҪПЈ»5) НЁ№эЦчіЙ·ЦН¶У°·ЦОц20О¬РОМ¬ҝХјдЦРөДұдТмДЈКҪЎЈ
<Ҫб№ыУлМЦВЫ>
Па№ШРФ·ЦОцПФКҫЛщУРІгҙОөДұдТмҫШХуҫщіКПФЦшХэПа№ШЎЈИзНј2ЛщКҫЈ¬·ўУэФлТфЈЁFЈ©УләкҪш»Ҝ·Ц»ҜЈЁRЈ©өД»Ш№йРұВКҰВ=0.67ЎА0.07ЧоОӘҪУҪьАнВЫФӨІвЦө1Ј¬ұнГч·ўУэПөНі¶ФіӨЖЪҪш»ҜДЈКҪөДЛЬФмЧчУГЎЈПаұИЦ®ПВЈ¬ТЕҙ«ұдТмЈЁGЈ©УлЦЦИә·Ц»ҜЈЁDpopЈ©өД№ШПөЛд·ыәП¶ЁБҝТЕҙ«АнВЫФӨІвЈЁҰВ=0.80ЎА0.16Ј©Ј¬ө«ҪвКНБҰПа¶ФУРПЮЎЈ

РОМ¬ұдТм·ЦОцҪТКҫұЈКШөДҝХјдДЈКҪЎЈИзНј3ЛщКҫЈ¬ЛщУРІгҙОөДұдТмЦчТӘјҜЦРФЪөЪ7-10әЕібВцұкЦҫөгөДІаПтО»ТЖЈ¬¶шіб°т»щІҝұкЦҫөгұдТмҪПРЎЎЈХвЦЦҝзІгҙОөДТ»ЦВРФ°өКҫ·ўУэПөНіҙжФЪДЪФЪөДұдТмЙъіЙЖ«ПтЈЁdevelopmental biasЈ©ЎЈ
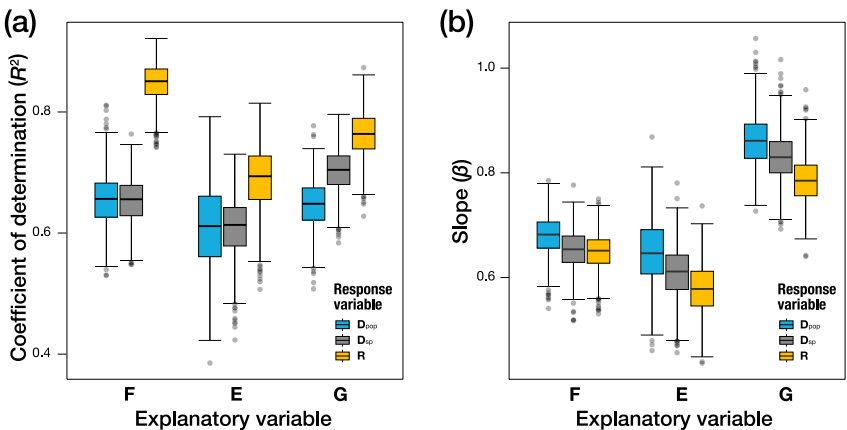
ұҙТ¶Л№әуСй·ЦОцСйЦӨБЛәЛРД·ўПЦөДОИҪЎРФЎЈИзНј4ЛщКҫЈ¬F-R№ШПөөДR2·ЦІјПФЦшёЯУЪЖдЛыЕд¶ФЈ¬ЗТІ»КЬРӯ·ҪІоҫШХу№АјЖОуІоУ°ПмЎЈСРҫҝХЯМбіцИэј¶ҙ«өј»ъЦЖЈә1) әкҪш»ҜАъК·ЛЬФм·ўУэПөНіІўҫц¶ЁұдТмРФЈЁF/MЈ©Ј»2) ·ўУэПөНіФЪұҫөШККУҰәНЖҜұдУ°ПмПВЛЬФмТЕҙ«ұдТмЈЁGЈ©әНҝЙЛЬРФЈЁEЈ©Ј»3) Gҫц¶ЁОўҪш»ҜДЈКҪЈЁDpopЈ©ЎЈ
ёГСРҫҝНЁ№эҪЁБўұдТм-·Ц»Ҝ№ШПөөДНіТ»ҝтјЬЈ¬ҪвҫцБЛҪш»ҜҝЙФӨІвРФгЈВЫЎЈІЙУГ"ұзЦӨ"КУҪЗЈЁdialectical perspectiveЈ©ХыәПІ»Н¬ЧйЦҜІгҙОЈ¬ұнГч·ўУэФлТфЧчОӘ"Ҫш»ҜІЁ¶Ҝ-ПмУҰ№ШПө"ЈЁevolutionary fluctuation-response relationshipЈ©өДМеПЦЈ¬ҝЙДЬКЗБ¬ҪУОўҪш»ҜУләкҪш»ҜөД№ШјьЕҰҙшЎЈХвР©·ўПЦІ»ҪцЦ§іЦSimpsonәНHansenөДҫӯөдәкҪш»ҜАнВЫЈ¬ТІОӘАнҪв·ўУэПөНіФЪҪш»ҜФӨІвЦРөДЧчУГМṩБЛРВЦӨҫЭЎЈСРҫҝҪб№ы·ўұнФЪЎ¶Communications BiologyЎ·Ј¬ОӘ·ўХ№ККУГУЪЛщУРКұјдіЯ¶ИөДНіТ»Ҫш»ҜАнВЫөм¶ЁБЛ»щҙЎЎЈ
ЙъОпНЁОўРЕ№«ЦЪәЕ
ЦӘГыЖуТөХРЖё
ҪсИХ¶ҜМ¬ | ИЛІЕКРіЎ | РВјјКхЧЁАё | ЦР№ъҝЖС§ИЛ | ФЖХ№МЁ | BioHot | ФЖҪІМГЦұІҘ | »бХ№ЦРРД | МШјЫЧЁАё | јјКхҝмС¶ | Гв·СКФУГ
°жИЁЛщУР ЙъОпНЁ
Copyright© eBiotrade.com, All Rights Reserved
БӘПөРЕПдЈә
ФБICPұё09063491әЕ
