
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
人类MRE11-RAD50-NBS1复合物感知DNA断裂的结构基础及其端粒因子TRF2调控机制
《Nature Communications》:Structural basis for DNA break sensing by human MRE11-RAD50-NBS1 and its regulation by telomeric factor TRF2
【字体: 大 中 小 】 时间:2025年09月20日 来源:Nature Communications 15.7
编辑推荐:
本研究针对DNA双链断裂(DSBs)检测与修复的核心机制,通过冷冻电镜技术解析了人源MRN复合物与DNA及端粒保护因子TRF2的结合结构,揭示了其通过闭合 coiled-coil 结构域形成钳状"感知状态"的DNA末端识别机制,发现NBS1的ATM招募基序被RAD50 S位点屏蔽的自动抑制状态,并阐明TRF2通过iDDR基序阻断S位点以抑制核酸酶活性和ATM激活的端粒保护机制,为理解哺乳动物MRN在DNA损伤应答中的功能分离提供了结构框架。
在基因组维护的精密调控网络中,DNA双链断裂(DSBs)是最致命的损伤类型之一,它们可能由电离辐射、基因毒性化学物质或复制叉崩溃自发产生,同时也是减数分裂重组和免疫球蛋白基因重排中的编程性中间体。若未能正确修复这些断裂,将导致染色体畸变、细胞死亡或癌变。细胞通过DNA损伤应答(DDR)机制应对DSBs,主要依赖ATM、ATR和DNA-PK激酶的激活,进而引发复杂的染色质修饰,并促进经典非同源末端连接(c-NHEJ)或同源重组(HR)修复途径。
在这一修复体系中,MRE11-RAD50-NBS1(MRN)复合物(在酵母中称为MRX)作为核心传感器,在DSBs的检测、信号传导和核酸酶处理中发挥多重功能。该复合物在进化上高度保守,由具有核酸酶活性的MRE11和ATP酶RAD50组成核心二聚体,并辅以真核生物特有的NBS1亚基。NBS1负责招募ATM激酶和辅助因子CtIP,而RAD50的coiled-coil结构域(CCDs)通过锌钩(zinc-hook)区域形成环状结构,充当ATP依赖的DNA末端识别门控机制。尽管前人对原核生物MR复合物的结构与功能已有深入研究,但人源MRN复合物如何感知普通DNA末端与端粒DNA,并实现末端感知与核酸酶活性的解耦,仍缺乏结构层面的精确阐释。此外,端粒保护因子TRF2(或酵母中的Rif2)可通过与RAD50的调控表面――S位点结合,抑制MRN依赖的ATM激活和核酸酶活性,然而这一调控的结构基础尚未明确。
为了解答这些关键问题,本研究团队通过冷冻电镜(cryo-EM)技术解析了人源MRN与DNA及TRF2复合物的高分辨率结构,系统揭示了MRN的DNA末端感知机制及其端粒特异性调控原理。相关研究成果于2025年9月18日发表在《Nature Communications》期刊。
研究团队主要运用以下关键技术方法:首先采用昆虫细胞和人类Expi293F细胞系统共表达并纯化人源MRN复合物及TRF2蛋白变体;通过荧光各向异性(FA)实验分析蛋白与DNA结合特性;利用NADH偶联的ATP酶活性测定评估MRN的酶活调控;采用单颗粒冷冻电镜技术解析了MRN与DNA(含ATP类似物)、MRN与TRF2-端粒DNA复合物的结构(最高分辨率达2.7?);结合化学交联质谱(CX-MS)验证复合物的相互作用网络;最后通过AlphaFold3预测的结构模型进行刚性对接和精细化建模。
研究人员发现人源MR/MRN复合物通过RAD50的核苷酸结合域(NBDs)以二重对称方式结合DNA,形成约20个碱基的足迹。DNA主要与RAD50的N-lobe和C-lobe相互作用,而MRE11在"感知状态"中几乎不直接接触DNA。特别值得注意的是,NBS1的C末端富含正电荷的序列(K751KRRR755)结合从RAD50伸出的DNA,增强了复合物的稳定性。结构分析显示RAD50的发夹环(1168-1183)插入DNA的小沟,同时两个CCDs向内倾斜,在DNA上方20?处相遇,形成紧密的钳状结构。
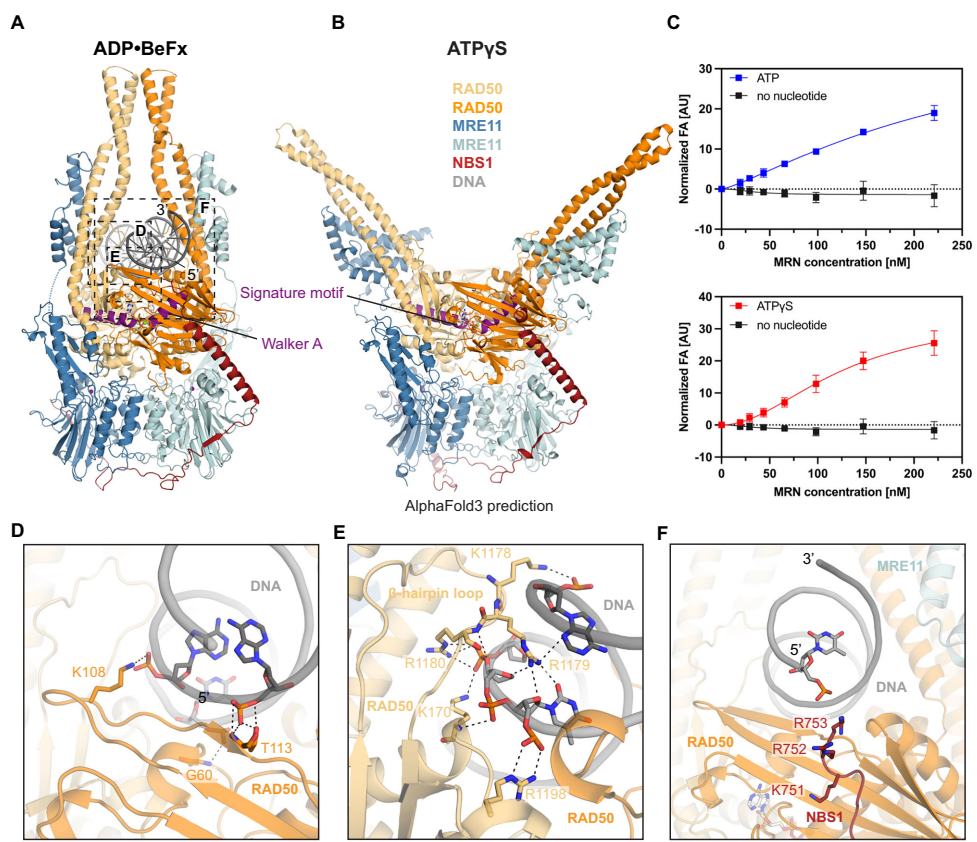
研究团队观察到MRN存在两种显著不同的构象状态:在ATPγS存在时,CCDs呈现开放状态,类似于原核MR的"静止状态";而在ATP水解后,CCDs闭合形成紧密的DNA钳制结构。令人惊讶的是,与人源MRN不同,闭合状态并未激活MRE11的核酸酶活性,而是保持自抑制状态。这种新发现的构象被命名为"感知状态",成功解释了真核生物中DNA末端识别与核酸酶活性的功能分离机制。
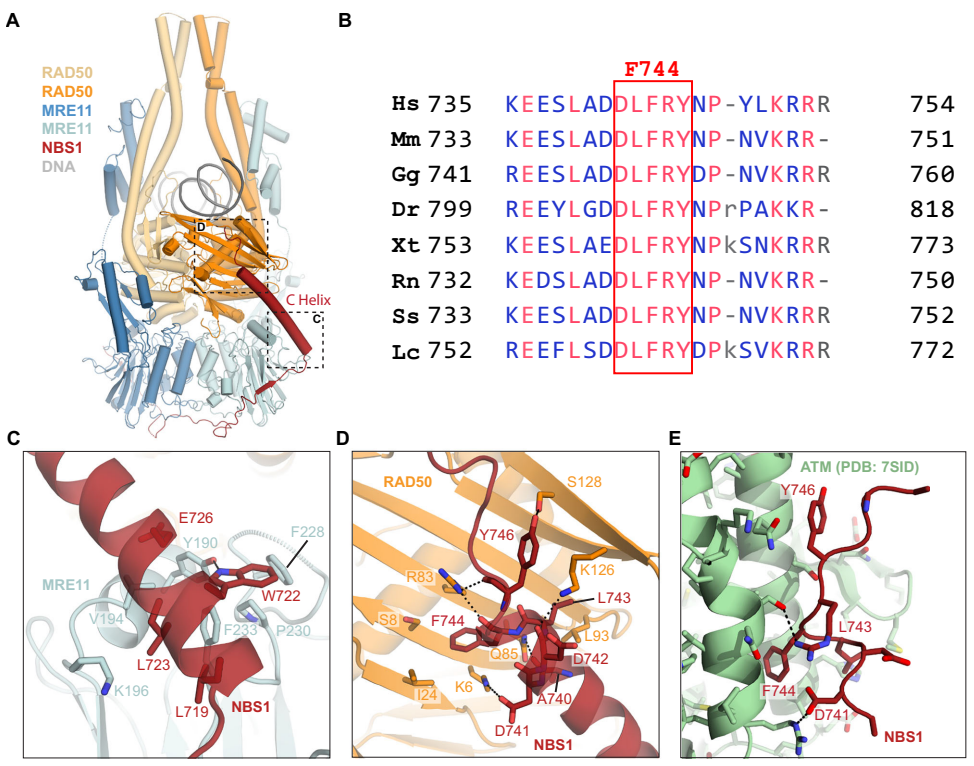
结构分析揭示NBS1(651-754)以单链形式包裹MRE11二聚体,其C端螺旋(718-745)同时结合MRE11催化结构域和RAD50的S位点。其中高度保守的F744嵌入由K6RAD50、S8RAD50等残基形成的疏水口袋。值得注意的是,NBS1的ATM招募基序(DLFRY序列)与S位点的结合模式与其与ATM的结合模式高度相似,表明MRN需要经历构象变化使NBS1 C螺旋从S位点解离才能激活ATM。
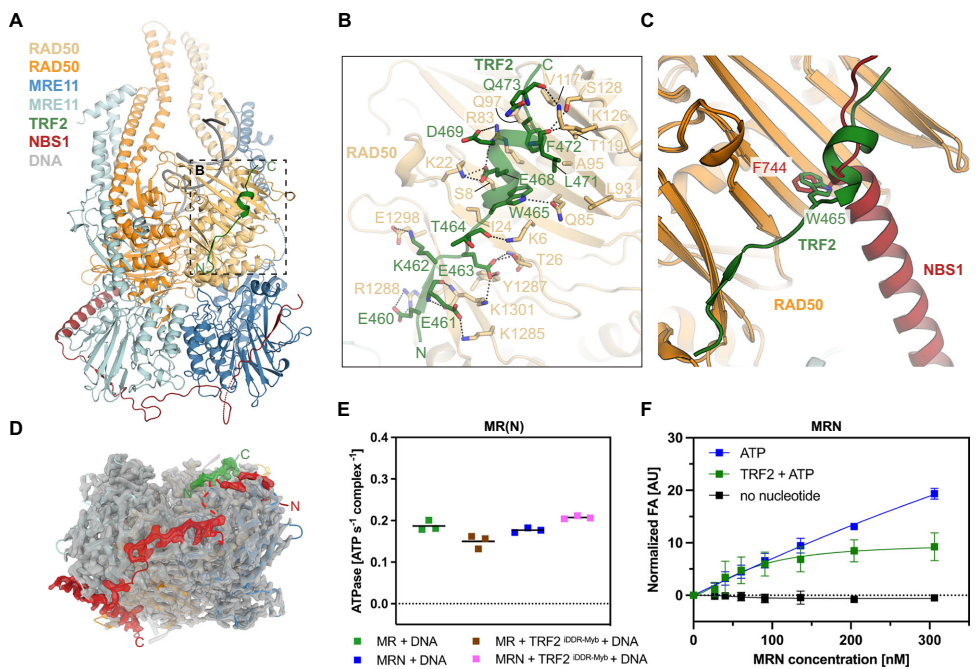
通过解析TRF2iDDR-Myb与MRN的复合物结构,研究人员发现TRF2的iDDR基序(459-473)以短螺旋形式结合RAD50的S位点,其中W465TRF2、L471TRF2和F472TRF2形成疏水锚定,酸性簇(E467EDE470TRF2)与RAD50的多个碱性残基产生静电相互作用。在MRN复合物中,TRF2仅能结合一个S位点,另一个位点已被NBS1的C螺旋占据,这种不对称结合实现了端粒处核酸酶活性的特异性抑制。
本研究系统揭示了人源MRN复合物感知DNA双链断裂的结构基础及其端粒特异性调控机制。研究发现的"感知状态"完美解释了真核生物中DNA末端识别与核酸酶活性的功能分离:MRN通过RAD50 CCDs的ATP依赖的环-杆转换机制加载到线性DNA末端,形成紧密钳制结构但保持核酸酶自抑制状态。这种机制使得MRN能够区分游离DNA末端和内部/环状DNA,为理解DNA损伤检测的拓扑学机制提供了重要见解。
特别重要的是,研究揭示了NBS1的C端螺旋与RAD50 S位点的相互作用机制,这为理解ATM激活的调控提供了结构基础:NBS1的ATM招募基序在感知状态中被S位点屏蔽,必须经过构象变化才能与ATM结合。这一发现解释了先前观察到的RAD50-NBS1直接相互作用,并为ATM激活需要MRN构象重排的观点提供了直接证据。
在端粒保护方面,研究阐明了TRF2通过iDDR基序结合S位点的精确分子机制,这种结合与NBS1的C螺旋结合模式既相似又竞争。值得注意的是,TRF2并不引起MRN从DNA上解离,反而可能通过稳定感知状态在功能失调的端粒处发挥结构性功能。这一发现修正了先前关于TRF2 simply去除MRN端粒的观点,揭示了更复杂的调控模式。
该研究的发现对理解癌症基因组不稳定性和端粒相关疾病具有重要意义,为开发针对MRN功能调控的新型治疗策略提供了结构基础。未来研究需要进一步阐明ATP水解在DNA末端处理中的精确作用,以及CtIP等辅助因子如何促进切割状态的形成等重要科学问题。
生物通微信公众号

