
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
稳定哺乳动物RNA温度计为蛛网膜下腔出血提供神经保护新策略
【字体: 大 中 小 】 时间:2025年09月20日 来源:Nature Communications 15.7
编辑推荐:
研究团队针对哺乳动物细胞如何感知微小温度变化调控基因表达这一核心问题,开展了RNA G-四链体(rG4)作为温度传感器的机制研究。发现低温或钾离子升高能稳定rG4结构,通过调控RBM3基因毒性外显子(poison exon)的可变剪接,促进神经保护蛋白RBM3表达。利用FDA批准的钾通道阻滞剂4-AP可模拟低温效应,在蛛网膜下腔出血小鼠模型中证实其通过RBM3依赖途径显著改善神经功能。该研究为治疗出血性脑卒中提供了全新靶点。
在哺乳动物中,核心体温被精确调控在37°C左右,但细胞如何感知和响应微小温度变化仍不完全清楚。虽然治疗性低温(therapeutic hypothermia, TH)是公认的神经保护策略,特别是在脑内和蛛网膜下腔出血(subarachnoid hemorrhage, SAH)中,但其分子机制和持续神经元保护策略仍不明确。近年来,冷休克蛋白RBM3(RNA-binding motif 3)被确定为TH神经保护作用的关键介质。在分子水平上,温度波动可能涉及哺乳动物的多种机制,甚至微小变化(-1°C)也会通过调节SR蛋白磷酸化影响RNA剪接。除了蛋白质介导的调控外,RNA本身由于其结构灵活性可能作为温度传感器。虽然RNA温度传感在细菌、酵母和植物中已有充分研究,但其在哺乳动物中对微小生理温度变化的响应作用仍 largely unexplored。
RNA G-四链体(rG4s)是由Hoogsteen鸟嘌呤碱基对(G-G)形成的相对稳定的二级结构。值得注意的是,富含G的RNA元件可以被G4配体(如PDS、Phen-DC3和钾离子)稳定,在体外和细胞中形成rG4s。它们在多种生物过程中发挥关键作用,参与可变多聚腺苷酸化、RNA颗粒的相分离和翻译等分子功能。此外,在哺乳动物细胞中的全基因组分析发现,剪接位点附近富含G的DNA元件和DNA G4结构。虽然G4元件在真核生物基因组中普遍存在,并在体外形成稳健结构,但关于rG4在哺乳动物细胞内的折叠仍存在 ongoing debate。重要的是,rG4作为生理动态、温度依赖性调控RNA结构在哺乳动物中的参与尚未被研究。
这项发表在《Nature Communications》的研究揭示了rG4作为哺乳动物RNA温度计的新机制,并通过稳定这些结构为蛛网膜下腔出血提供了新的治疗策略。
研究人员采用RNA测序、生物信息学分析、分子生物学实验和生物物理技术相结合的方法,利用HEK293T、Hela和HT22细胞系以及SAH小鼠模型,通过体内外实验验证了rG4的温度感应功能及其神经保护机制。人类样本未涉及本研究。
研究发现rG4结构非常适合作为哺乳动物细胞中的生理温度传感器。通过分析不同温度下的RNA-seq数据,计算 cassette外显子剪接位点周围序列的G含量,将外显子分为低温抑制外显子(CREs)、非温度敏感外显子(NTs)和低温诱导外显子(CIEs)。发现在温度敏感外显子中,侧翼序列的G含量显著高于CIEs和NTs。这种差异在上游和下游组成型外显子的剪接位点侧翼序列中未观察到,表明高G含量是低温抑制型cassette外显子的特异性特征。

在不同细胞模型中的验证显示,在冷休克(32°C vs 37°C)时,CREs剪接位点侧翼序列的G含量显著高于CIEs和NTs。然而,在热休克(40°C vs 37°C)时未观察到这种富集,表明富含G的序列可能在较低温度下优先发挥作用。通过搜索先前研究中的motif,确实观察到CREs的5'-剪接位点侧翼序列中rG4 motif的强烈富集。使用G4Hunter扫描进一步证实了CREs的G4评分高于CIEs和NTs。分析rG4-seq数据发现,在rG4稳定剂PDS处理的细胞中,rG4-seq峰在剪接位点周围富集,特别是在CREs的3'和5'剪接位点侧翼强烈富集rG4s。
为了探讨rG4在控制温度依赖性可变剪接中的全局作用,研究人员将预测的G4评分与冷或热休克处理后外显子包含水平的变化(delta PSI)相关联。发现温度敏感外显子5'-剪接位点周围的预测G4评分与低温下外显子排除增加相关,而热休克时观察到相反但稍弱的相关性。这一结果表明剪接位点周围rG4的折叠概率(预测G4评分)与低温下外显子排除之间存在普遍相关性。
进一步重叠Hela和HEK293T中鉴定的CREs,获得一组高置信度外显子,排除细胞类型特异性调控的外显子。总共1065个常见CREs被鉴定,其中380个(35.7%)在其剪接位点周围含有预测的rG4s(评分>1),指向一种普遍的、细胞类型独立的剪接调控功能。使用三种代表性基因(CDK4、IQSEC1和FKBP15)作为对照,这些基因在控制细胞周期、磷酸肌醇代谢和细胞骨架组织中具有 distinct功能。序列比对显示CREs剪接位点周围的G4在哺乳动物中保守,表明它们在进化中的重要性。
通过用广泛使用的G4稳定剂PDS处理HEK293T和Hela细胞,在不同温度(33、35、37和40°C)下测量这些CREs的包含水平。分析显示CREs对PDS的存在具有强烈敏感性,导致在较高温度下外显子排除,初步表明rG4稳定化抑制外显子包含。还使用两种额外的G4配体(Phen-DC3和NMM)在HEK293细胞中评估配体限制效应,并测试了几个CREs(CDK4、IQSEC1和RBM3)。Phen-DC3处理显著减少了这些外显子的包含,而NMM没有效果,可能由于不同稳定剂对不同rG4结构的差异响应。
接下来研究人员进一步证实富含G元件与温度敏感性可变剪接之间的联系。聚焦于冷诱导和神经保护蛋白RBM3,其通过温度调控的可变剪接控制一个毒性外显子(外显子3a),导致热诱导的NMD。在RNA-Seq数据中,RBM3的外显子3a包含水平在37°C或以下始终低于0.1,而在39°C或40°C时显著增加。在hRBM3和mRBM3微型基因中,外显子3a的剪接保持温度敏感性,G4配体PDS几乎完全阻断所有测试温度下的外显子3a包含。此外,在这个冷抑制毒性外显子的3'和5'-剪接位点周围鉴定了推定的rG4。
为了探索RBM3外显子3a剪接位点附近富含G元件的影响,在五个区域引入了突变:两个围绕剪接位点(R1和R3),一个在外显子3a内(R2),两个在下游内含子中(R4和R5)。有趣的是,R1和R3中富含G元件的突变或缺失增加了HEK293T和小鼠N2a细胞中的外显子3a包含。使用MaxEntScan进一步排除了R1突变促进外显子3a包含是由于增加3'-剪接位点强度的可能性。
总之,诱变分析支持这样的假设:R1和R3区域中的富含G元件通过掩盖紧密相邻的剪接位点或招募剪接调控蛋白,促进RBM3外显子3a可变剪接。R1和R3中四个连续G stretches可能形成rG4结构,在哺乳动物中进化保守,表明它们在调控RBM3表达中的重要性。R13-2G双突变体在HEK293T细胞和N2a细胞中增加了包含外显子3a的异构体,效果比每个单突变体更强,再次证实这些剪接位点附近的富含G序列抑制外显子3a包含。使用RBM3 R13-2G微型基因的进一步分析显示,它对G4配体失去响应性,确认它们通过预测的G4结构起作用。
为了进一步验证富含G元件R1和R3对可变剪接的温度敏感性抑制效应,研究人员将R1和/或R3 motif引入一个无关的微型基因(MINX或荧光报告基因),并比较了WT motif与其突变或对照对应物对可变剪接的影响。这种方法使我们能够确定富含G motif是否在无关分子 setting 中在细胞背景下赋予温度敏感性可变剪接。在MINX微型基因测定中,将RBM3 3a外显子的一段,包括WT或突变的富含G元件, incorporated 在MINX基因的3'剪接位点。WT序列在较低温度下表现出增加的外显子跳跃,与突变体相比,在升高温度下跳跃减少,支持这个富含G R1元件作为哺乳动物细胞中调控可变剪接的温度传感器的作用。此外,使用荧光微型基因报告系统,其中cassette外显子的包含破坏DsRed阅读框,阻止其表达同时允许GFP翻译,而外显子跳跃维持DsRed框架,带有终止密码子在GFP翻译前终止翻译。在cassette外显子剪接位点附近引入R1和/或R3显著增加DsRed信号同时减少GFP信号,导致更高的DsRed/GFP比率,表明R1和/或R3引起的外显子跳跃增加。值得注意的是,抑制效应在39°C减少,突出了富含G元件介导的剪接调控的温度依赖性特征。重要的是,这个微型基因仅在剪接位点附近存在富含G motif时表现出温度响应性,支持这些结构作为温度传感器调控可变剪接的假设。
研究结果证明围绕RBM3外显子3a的富含G元件通过限制剪接位点可及性在较低温度下调控其包含。为了提供这些序列形成rG4结构并表现出温度敏感性的直接证据,使用了对应于R1序列(rG4-R1)的RNA寡核苷酸和一个突变版本(R1突变体),其中关键G残基被替换为Cs。圆二色谱(CD)光谱学,一种检测rG4的标准方法,显示R1 RNA在265 nm处有强正峰和在240 nm处有负峰, characteristic 平行rG4结构。这个信号依赖于KCl,一种内源性G4稳定剂,在R1突变体中缺失,确认体外rG4形成。进一步分析显示在低钾浓度(0.1mM)下,rG4形成是温度敏感的,当加热到40°C并随后冷却到33°C时信号强度可逆变化。相比之下,在高钾浓度(50mM)下,温度敏感性降低。
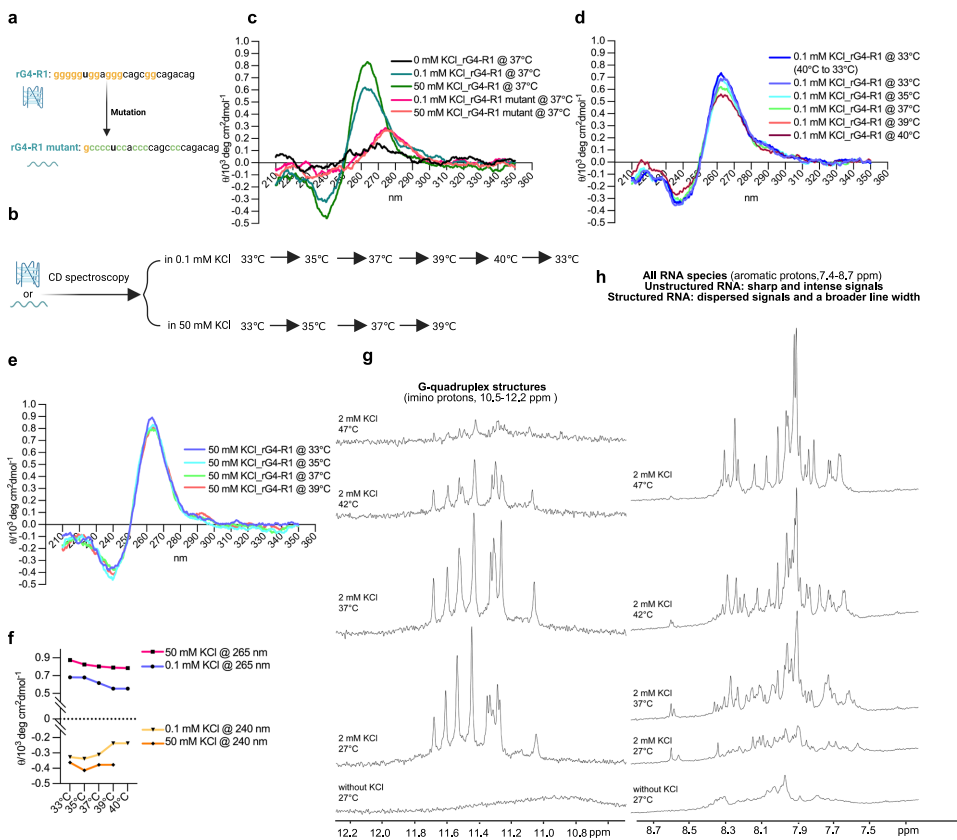
为了进一步验证rG4形成,记录了在不同温度和钾浓度下rG4-R1 RNA的1D 1H NMR光谱。亚氨基质子区域(10-16 ppm)指示RNA二级结构,Watson-Crick碱基配对信号出现在12-15ppm,rG4特异性信号在10.5-12.2 ppm的狭窄范围内。在没有钾的情况下,未检测到亚氨基信号,表明缺乏稳定的RNA结构。 upon 钾添加,在27°C时良好分散的亚氨基信号确认rG4形成。温度依赖性NMR分析显示rG4信号在37°C和42°C之间减弱,表明部分rG4 destabilization,在47°C时几乎不可检测。在芳香质子区域(7.4-8.7 ppm),非结构化RNA的信号在27°C时缺失,但在37、42和47°C时显著增加,表明在较高温度下逐步rG4展开。值得注意的是,体外CD和NMR中使用的钾浓度不直接反映细胞条件,其中额外因素 contribute to rG4稳定性。例如,RNA解旋酶(如DHX36)和RNA结合蛋白(如hnRNPF)可以溶解rG4s,这使得rG4s在细胞环境中即使在生理钾浓度140-150 mM下也不那么稳定。
总之,这些结果强烈支持形成温度响应性rG4结构,动态响应生理温度范围内的变化。这些发现共同确立rG4作为哺乳动物RNA温度计,能够调控温度依赖性可变剪接。
为了进一步研究不同温度下活细胞中rG4的折叠状态,进行了G4特异性免疫染色和SHAPE-MaP探测,以直接评估rG4折叠动力学。细胞暴露于低温(33°C)显著增加G4特异性信号,这被benzonase(一种核酸酶) largely 废除,但仅被DNase处理部分减少,主要影响核染色。这支持rG4折叠在低温下在活细胞中 promoted 的观点,并且RNA,主要在细胞质中检测到,能够形成rG4。此外,检查扰动是否影响G4信号,使用电压门控钾通道阻滞剂(AFP, Amifampridine)在谷氨酸去极化/兴奋的HT22细胞中增加细胞内钾,发现AFP也显著增加这些细胞中的全局G4信号,表明增加细胞内钾促进rG4形成。
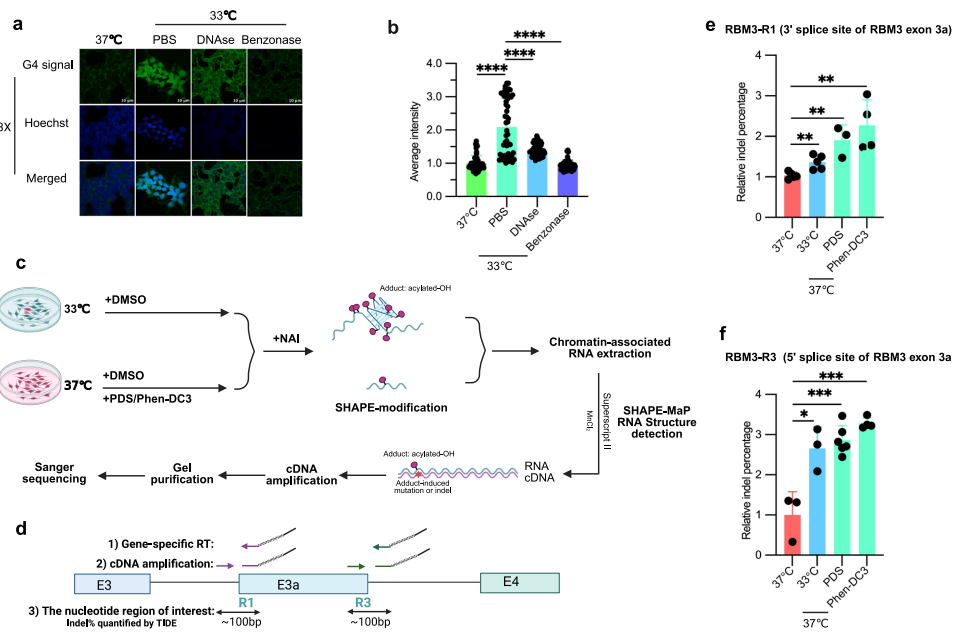
为了进一步证实rG4作为生理温度传感器调控哺乳动物细胞可变剪接的作用,进行了SHAPE-MaP探测。在这种方法中,NAI(2-methylnicotinic acid imidazolide)优先修饰RNA核糖糖柔性区域中的2'-羟基,导致连接G-四联体的环核苷酸、G4结构内的凸出残基以及相邻非结构化区域的更高修饰,而具有稳定碱基配对的区域和G-四联体堆叠区域将显示减少的修饰。NAI诱导的修饰将在逆转录期间导致cDNA中的突变或indels,使用Mn2+和Superscript II逆转录酶,可以通过TIDE软件量化。结果表明,在活细胞中用G4配体(PDS和Phen-DC3)处理显著增加几个冷抑制外显子(RBM3、CDK4和FKBP15)剪接位点附近富含G motif周围的indel百分比,表明这些motif可以在活细胞中形成rG4结构。更重要的是,indel百分比在低温(33°C)下显著增加,支持这些富含G元件在冷条件下形成更多rG4s。总之,这些发现提供了强有力的证据,表明rG4作为生理钾和温度传感器发挥作用,然后可以调控下游加工事件。
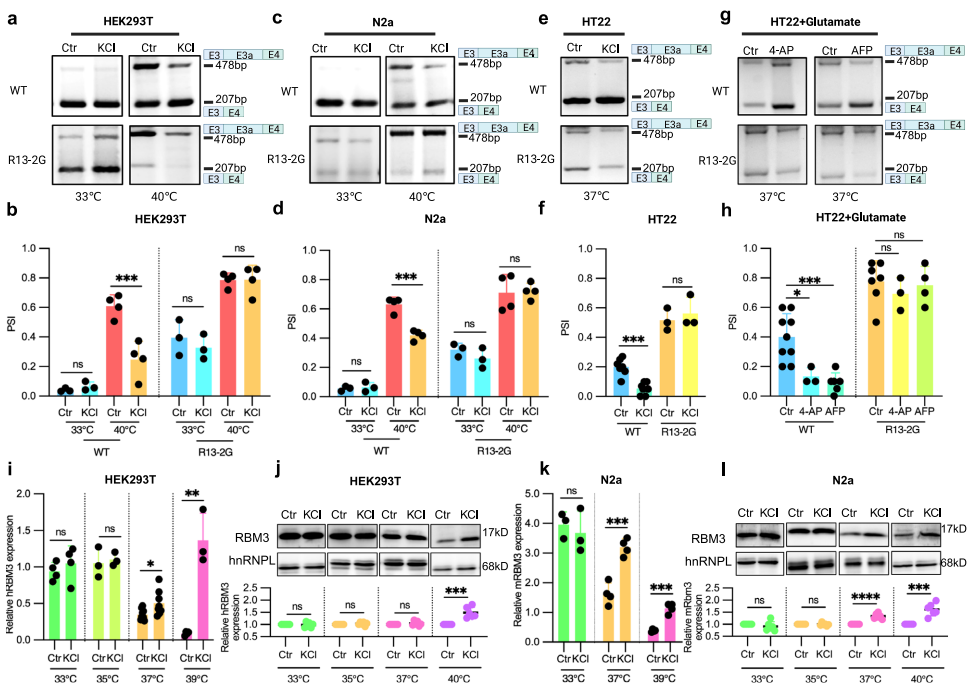
在证明RBM3外显子3a剪接位点周围的富含G序列可以在体外和体内形成rG4结构后,研究人员旨在确认它们对细胞中RBM3外显子3a剪接的相关性。因此,转向内源性rG4稳定剂钾(K+)并测试KCl对mRBM3微型基因和rG4双突变体(R13-2G)在HEK293T和N2a细胞中的影响。在WT mRBM3微型基因中,KCl处理在高温下显著减少RBM3外显子3a包含,而在低温下没有效果。这表明在较低温度下rG4结构的更高稳定性导致对钾的依赖性降低,在高温下,钾稳定rG4,导致更多RBM3外显子3a跳跃。相比之下,双突变体(R13-2G)中的外显子3a包含在低温和高温下 upon KCl处理没有显著变化。一致地,KCl在正常生理温度37°C下在HT22细胞中促进WT mRBM3微型基因中的外显子3a跳跃,而对R13-2G突变微型基因没有影响。在极化/谷氨酸激活的HT22细胞中,模拟病理性脑损伤条件,4-AP和AFP都显著减少WT中的外显子3a包含,但不在R13-2G微型基因中。这些结果共同强烈表明RBM3外显子3a中的rG4是KCl响应性和温度依赖性RNA元件,通过掩盖剪接位点,控制细胞中的外显子包含。
由于RBM3外显子3a包含触发无义介导的衰变(NMD),通过使用钾稳定其剪接位点附近的rG4促进外显子3a跳跃可以增强内源性RBM3表达。因此,使用KCl稳定RBM3外显子3a中的rG4,并研究其对内源性RBM3表达的影响。与假设一致,KCl处理在高温(≥37°C)下增加RBM3 mRNA和蛋白质表达,但在低温(<37°C)下没有。总之,这些发现表明低温和钾离子都能够稳定rG4结构,从而增加内源性RBM3表达。
由于已经 well established RBM3保护神经元并在朊病毒疾病和缺氧缺血性脑损伤小鼠模型中通过促进防止细胞死亡和促进神经发生改善表型,研究人员随后调查了通过增加细胞内钾浓度与4-AP(一种临床批准的化合物)稳定rG4结构是否可以在血红素诱导的出血细胞模型中提供神经保护作用。在这个模型中,血红素,一种血红素衍生物――红细胞中血红蛋白的基本组成部分――模拟出血性卒中早期脑损伤的氧化应激和神经炎症。观察到血红素处理导致细胞内钾浓度显著减少,而4-AP处理在血红素诱导的细胞培养模型中产生剂量依赖性的细胞内钾增加。接下来首先使用三种不同的引物对测量RBM3外显子3a的剪接水平(引物对1用于3'剪接位点连接,对2用于5'剪接位点连接)。血红素处理显著增加外显子3a包含,通过外显子3a与RBM3 mRNA的更高比率证明, ultimately 导致RBM3表达几乎两倍减少,而4-AP处理导致外显子3a与总RBM3比率的逐渐减少和RBM3 mRNA表达的增加。因此,RBM3蛋白质表达在4-AP处理后 substantially 增加,与单独血红素处理相比达到 nearly 五倍升高。表型上,血红素诱导的细胞死亡率减少,从25%到5%,细胞活力在4-AP处理后显著和 substantially 增加。用siRNA敲低RBM3后4-AP处理废除了这些表型效应,强调了RBM3在介导4-AP赋予的神经保护中的关键作用。为了进一步验证RBM3对对抗血红素诱导的神经元损伤的保护作用,在HT22细胞中过表达RBM3。这导致细胞活力从60%增加到80%,并在血红素处理时细胞死亡显著下降从16%到7%。

为了进一步解决4-AP的功效和临床应用潜力,研究人员随后使用蛛网膜下腔出血(SAH)小鼠模型评估了4-AP的影响。小鼠的4-AP剂量基于先前建立的方案,以确保有效性同时最小化显著副作用。鉴于SAH个体中最受影响区域 near 出血部位,关注受影响区域周围的大脑皮层。首先检查了4-AP给药后的RBM3表达水平。腹腔注射4-AP导致大脑皮层中细胞内钾水平显著升高,内源性RBM3 mRNA表达上调,RBM3外显子3a包含减少。RBM3蛋白质表达也增强,如免疫染色所示。接下来,分析神经元凋亡、神经元计数和皮层海绵状作为关键读数,以评估4-AP在SAH模型中的效果。值得注意的是,4-AP处理后增加的RBM3表达与减少的神经元凋亡和增加的神经元计数相关。苏木精和伊红(HE)染色也显示4-AP处理显著改善SAH导致的皮层海绵状严重程度。4-AP给药还防止神经元损伤,表现为嗜酸性坏死神经元 with 异常形态特征,包括细胞体收缩、深染固缩核和强烈染色嗜酸性,神经元计数从 around 20显著增加到100。此外,Nissl染色显示与SAH对照组相比,4-AP处理后细胞计数增加 with 正常形态。最后,由于SAH模型在Willis环附近诱导出血,由于蛛网膜下腔血液积累和继发性缺血导致急性神经功能缺损,使用rotarod测试评估运动功能,这是啮齿动物协调和平衡的标准测量,并使用修改的Garcia评分评估整体神经功能缺损。4-AP处理显著改善小鼠行为表现,表现为增加的修改Garcia评分和rotarod测试中增加的跌倒潜伏期。
为了确认4-AP处理在SAH小鼠模型中的有益效果是否依赖于增加的RBM3表达,使用shRNA慢病毒在SAH前敲低体内RBM3。RBM3敲低废除了4-AP赋予的神经保护,使用先前建立的神经元读数和行为评估。因此,这导致增加的神经元凋亡,减少的神经元计数, elevated 海绵状评分,和减少的健康神经元,与单独4-AP处理相比。此外,4-AP处理引起的小鼠行为改善被RBM3敲低无效,表现为减少的修改Garcia评分和rotarod测试中减少的跌倒潜伏期。总之,这些结果强烈支持RBM3作为4-AP神经元效果的关键介质的作用。
为了进一步说明RBM3对SAH的神经保护功效,通过慢病毒介导的RBM3过表达在SAH前在体内增强小鼠中的RBM3表达。增加的RBM3水平与减少的海绵状严重程度、增加的健康神经元和增加具有正常形态的神经元计数显著相关。RBM3上调还显著提高SAH小鼠的Garcia评分,并在rotarod测试中适度改善表现,表现为增加的跌倒潜伏期。
总之,研究结果表明剪接位点附近的rG4可以作为哺乳动物温度传感器调控可变剪接网络。具体来说,RBM3外显子3a剪接位点周围的富含G序列在低温(33°C和35°C)或高钾浓度下形成稳定的rG4结构,抑制外显子包含,通过防止NMD导致神经保护性
生物通微信公众号
知名企业招聘

