
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
发育过程中上肢肌肉协同的精细化分馏:学龄前与学龄期儿童投掷动作的神经肌肉机制研究
【字体: 大 中 小 】 时间:2025年09月20日 来源:iScience 4.1
编辑推荐:
本刊推荐:为揭示儿童投掷技能发展的神经肌肉机制,研究人员开展了学龄前(3-5岁)与学龄期(6-9岁)儿童上肢肌肉协同(Muscle Synergies)的对比研究。通过肌电信号分析和非负矩阵分解(NMF)技术,发现学龄期儿童表现出更高的肌肉协同稀疏性(Sparseness)和协同簇分馏(Fractionation)现象,表明其肌肉招募更具选择性。该研究揭示了通过协同分馏实现的神经适应性优化是运动功能发展的重要机制,为理解人类运动控制的发育 plasticity 提供了新视角。
人类投掷能力深深植根于进化史,早期人类依靠投掷投射物来获取食物和保护资源。虽然现代社会对这种生存技能的依赖减少,但投掷作为一项基础运动行为,在棒球、板球和网球等运动中仍然至关重要。这种能力被归类为一种先天的运动行为,在儿童早期出现并在发育过程中逐步改善,即使没有特定训练也是如此。
然而,投掷表现的精进需要许多肌肉的协调和顺序激活,其模式在发育过程中受多种因素影响。中枢神经系统(CNS)通过不断调整运动指令,以适应神经肌肉骨骼系统的变化(如肌肉力量、刚度和神经运动反射增益)和任务复杂性(需要精确计时、力生成和肌肉激活顺序)。研究表明,CNS可能依赖一种模块化架构,即运动原始模块或运动模块,来协调和执行涉及肢体数千个运动单位的各种运动行为。一些运动模块的结构可能是天生的或生命早期确定的,并在大部分寿命中保持相对不变,但也有研究表明,模块会被新生成或修改以满足新复杂技能的机械需求。
为了探索发育过程中投掷运动的神经可塑性,研究人员重点关注了投掷作为基础运动预计会显著改善的发展阶段。特别是,在幼儿期如学龄前儿童(3-6岁),儿童表现出较低的功能技能水平,而投掷表现(如球速度和准确性)在学龄期(6-9岁)显著提高。这种改善可能与神经肌肉控制的适应性变化有关,但具体的神经机制尚不清楚。
为了解决这一问题,研究人员开展了了一项研究,比较学龄前和学龄期儿童在过顶投掷过程中的上肢肌肉协同。该研究旨在揭示发育过程中肌肉协同结构的变化,以及这些变化如何与投掷表现的改善相关。研究结果发表在《iScience》期刊上,为理解人类运动控制的发育 plasticity 提供了新的见解。
研究人员主要应用了肌电图(EMG)技术记录了16块上肢肌肉的活动,并结合高速摄像系统捕捉投掷动作的球体运动学数据。通过非负矩阵分解(NMF)方法从EMG数据中提取肌肉协同,分析了协同的结构和稀疏性。此外,采用层次聚类分析确定了代表性协同簇,并通过统计参数映射(SPM)比较了时间模式的组间差异。研究还计算了协同稀疏性指标,以评估肌肉招募的选择性,并利用非负最小二乘法模型分析了协同分馏现象。样本来源于21名健康儿童(13名学龄前儿童和8名学龄期儿童),所有参与者均完成了针对固定目标的投掷任务。
学龄期儿童在年龄、身高、体重和功能技能(通过PEDI-FSS-SC1评估)方面均显著高于学龄前儿童。更重要的是,学龄期儿童表现出更优的投掷表现:球速更高(6.14 m/s vs. 4.72 m/s),准确性更好,体现在更小的垂直误差(0.04 m vs. 0.23 m)和绝对距离误差(0.18 m vs. 0.43 m)。然而,投掷动作的时序参数(如运动开始到加速开始的时间、加速开始到球释放的时间)在两组之间没有显著差异。三维球轨迹显示学龄期儿童有更大的垂直和侧向运动。
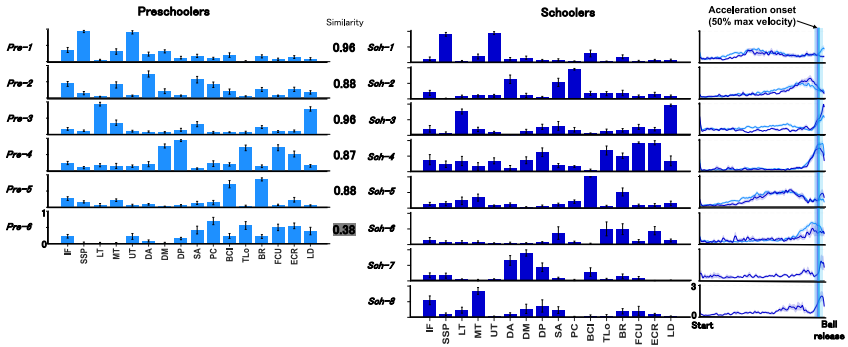
通过NMF提取肌肉协同后,发现两组在协同数量上没有显著差异(学龄前儿童中位数6,学龄期儿童中位数6.5)。层次聚类分析揭示了更复杂的协同组织:学龄期儿童有8个协同簇,而学龄前儿童只有6个。其中,5个协同簇在两组之间共享(标量积SP > 0.75),表明存在共同的神经肌肉协调策略。此外,学龄期儿童有1个独特协同簇,学龄前儿童有3个独特协同簇。这些共享协同的时序模式通过SPM分析没有显示组间显著差异。
协同稀疏性(Sparseness)在学龄期儿童中显著更高(0.53 vs. 0.50),表明其肌肉招募更加选择性和功能特异性,而学龄前儿童表现出更广泛的共激活。进一步分析表明,这种稀疏性增加可以通过协同分馏(Fractionation)解释:学龄前儿童的多个协同(Pre-2、Pre-4、Pre-5、Pre-6)在学龄期儿童中分馏成两个或多个功能特化的协同。例如,Pre-2(涉及肩关节推动和稳定肌肉)分馏为Sch-2(主导推动动作)和Sch-8(主导肩外旋),Pre-4分馏为Sch-4(主导肘和腕控制)和Sch-7(主导特定稳定功能)。

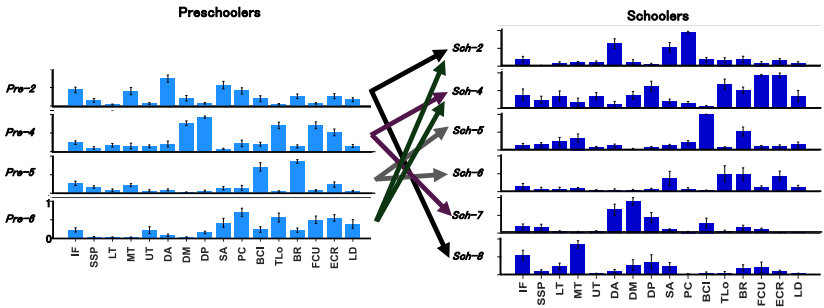
肌肉协同由分布在中枢神经系统(CNS)多层次的复杂神经网络实例化,包括皮质运动区、脑干核和脊髓前角中间神经元。在发育早期,由于感觉和下行调制不成熟,CNS主要依赖脊髓和脑干电路提供原始协调。随着儿童成长,神经肌肉骨骼特性的成熟增强了感觉信号,皮质脊髓束的成熟(包括髓鞘化和突触 refinement)使得对原始协同内的肌肉亚群进行区分控制成为可能。协同分馏的可能机制是CNS招募额外的神经电路来细分控制:如果先前 inactive 的脊髓电路变得可用,下行通路可以重新利用该电路,将其分配给控制先前由另一个协同共激活的肌肉子集。通过重复激活和差异化计时信号,这些新的神经连接得到加强。
这种 refinements 在功能上增强了投掷表现:更稀疏和分馏的协同允许更协调的投掷序列,减少关节共激活,增强能量传递效率和释放控制。例如,Sch-8 中肩外旋肌肉的专门化允许更长的加速阶段,更好地控制球释放和轨迹;Sch-2 中推动肌肉的巩固优化了动能传递。这些变化与先前研究一致,表明发育涉及从简单的、同步化的关节协调模式向更具多样性和时序变异性的模式转变。
该研究揭示了从学龄前到学龄期儿童投掷运动中上肢肌肉协同的显著可塑性。学龄期儿童表现出更高的投掷速度、准确性(特别是垂直方向)和更大的肌肉协同稀疏性,反映了更选择性的肌肉招募。协同分馏是关键的发育适应机制:多个学龄前儿童协同分馏成学龄期儿童中功能特化的组件,从而减少了共激活,增强了能量传递效率和释放控制。尽管一些基础协同在两组之间共享(表明早期出现和保存),但增加的模块复杂性为CNS提供了更丰富的运动组件“工具箱”,可以灵活组合和适应。
这些发现强调了 fractionation 驱动的专门化作为支持改善运动性能的关键发育适应。从临床角度来看,较少分馏的、共激活的协同模式(如年轻儿童或神经系统损伤患者中所见)会限制性能,而增加模块复杂性有利于运动输出。未来研究应调查 specifically 促进模块复杂性或最佳协同分馏的干预措施。
研究的主要限制在于缺乏详细的手臂运动学数据,这限制了对运动阶段(如 cocking phase)的精确定义和肌肉协同激活的机械解释。此外,尽管目标高度在年龄组之间标准化,但参与者身高的固有差异可能影响了投掷模式和协同招募。
生物通微信公众号
知名企业招聘

