
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
综述:偏头痛治疗进展
《TRENDS IN Molecular Medicine》:Migraine: advances in treatment
【字体: 大 中 小 】 时间:2025年09月20日 来源:TRENDS IN Molecular Medicine 13.8
编辑推荐:
本综述系统梳理了偏头痛治疗领域的最新突破,重点介绍了靶向降钙素基因相关肽(CGRP)及其受体、5HT1F受体(ditans)的创新疗法,并探讨了针对垂体腺苷酸环化酶激活多肽(PACAP)、肾上腺髓质素、胰淀素及其受体,以及KATP通道、瞬时受体电位(TRP)离子通道和神经元型一氧化氮合酶(nNOS)等新兴靶点的治疗潜力。文章强调患者异质性是实现精准医疗的关键。
偏头痛是一种复发性原发性头痛疾病,其特点是未经治疗时头痛持续超过4小时,并至少具有以下两种特征:单侧定位、搏动性、中度至重度强度、身体活动会加剧或导致回避活动,以及恶心或畏光和畏声。过去几十年,偏头痛的急性和预防性治疗均出现了新型疗法,例如曲普坦类(5HT1B/1D受体激动剂)、gepants(降钙素基因相关肽拮抗剂)和ditans(5HT1F受体激动剂)。针对CGRP机制的新特异性预防疗法已被确定。然而,偏头痛仍然非常普遍,在使用残疾损失健康生命年这一指标衡量时,其在所有健康状况中排名第三。尽管有可用疗法,仍有相当比例患者承受着巨大的疾病负担,约三分之一患者对CGRP单克隆抗体(mAbs)无应答,因此仍需进一步开发治疗选择。
鉴于偏头痛作为一种疾病具有多阶段性,许多患者除了头痛外还有前驱症状和头痛后期症状,并且这些非头痛症状(包括泌尿或胃肠道紊乱、食物渴望和情绪障碍)范围广泛且存在个体间差异,偏头痛的潜在病理生理学可能反映了一种神经网络障碍,而非 primarily 血管性障碍或单一解剖基底的功能障碍。三叉颈复合体被认为是偏头痛症状学的关键,是投射到硬脑膜纤维(导致疼痛感)的外周传出神经与中枢神经系统(如脊髓三叉神经核、上颈髓,然后上行至脑干、下丘脑和丘脑的第二级神经元)之间连接的枢纽。这些后述通路许多是双向的,例如下丘脑已被证明可调节三叉神经血管系统中的Sp5c神经元。尽管偏头痛生物学的起源仍是一个激烈辩论的话题,但与这些偏头痛结构和通路相关的分子靶点已被识别,从而推动了更新型偏头痛疗法的开发。
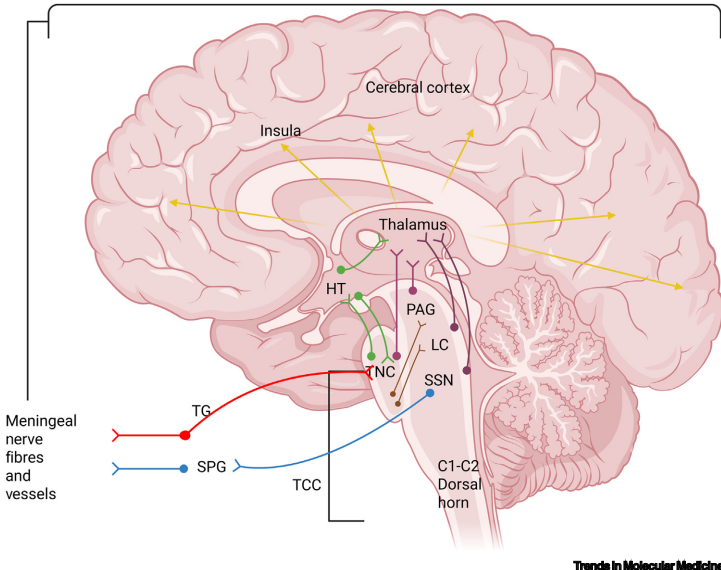
CGRP最初被鉴定为降钙素基因产生的RNA经过组织特异性加工的产物,并被表征为脑膜血管系统的有效血管扩张剂。后续研究在动物和人类模型中证明,激活三叉神经血管系统会释放CGRP,并且在偏头痛发作期间血清CGRP水平升高。CGRP在神经系统中广泛表达,包括偏头痛关键结构,如三叉神经核、外周伤害性脑膜纤维、下丘脑、颈背根神经节、丘脑和脑干。输注CGRP已被证明可在偏头痛患者中引发偏头痛样发作,进一步验证了CGRP在偏头痛生物学中的重要性。
钙素受体样受体(CLR)与受体活性修饰蛋白1(RAMP1)形成异源二聚体受体,构成CGRP受体。有证据表明CGRP受体组分蛋白(RCP)也是G蛋白偶联信号转导所必需的,一项研究表明,反义RCP构建体即使在不影响CGRP与受体结合的情况下,也能减弱CGRP信号转导。这些受体在整个大脑中也广泛表达,因此CGRP受体拮抗剂的确切作用部位尚未阐明,或者可能存在多个提供治疗益处的作用部位。CGRP也存在于颅血管的平滑肌细胞中,在那里它已被证明具有有效的血管舒张作用,这解释了为什么在偏头痛患者和疾病模型中观察到颅血管舒张的附带现象。重要的是要考虑到,虽然CGRP在三叉神经血管系统中的释放在偏头痛病理生理学中起关键作用已是公认事实,但这种释放的原因尚未完全确定。除了通过抗体拮抗CGRP外,DNA甲基化可能是靶向该通路的另一途径。
为靶向CGRP通路而创建的第一类药物是gepants,即针对CGRP肽的小分子抗体。CGRP在神经系统外的组织中也有表达,因此存在脱靶效应或代谢物毒性这类通用问题的理论风险。第一代gepants证明了临床疗效,但因肝酶升高和潜在肝毒性而停用。一个名为DILISym的定量系统毒理学模型用于一项试验,证明第一代gepants替卡吉泮(telcagepant)的肝酶升高可进展为药物性肝损伤(DILI),主要继发于对胆汁酸转运蛋白的抑制。另一个第一代gepants奥塞吉泮(olcegepant)是静脉给药,因正在开发口服生物可利用的替代品而停用。
进一步开发的gepants是口服给药,被称为“第二代”:瑞美吉泮(rimegepant)、乌布吉泮(ubrogepant)和阿托吉泮(atogepant)。瑞美吉泮和乌布吉泮在临床试验中证明了用于偏头痛急性治疗的功效,而阿托吉泮在每日使用时证明了作为偏头痛预防性药物的功效。一项关于扎维吉泮(zavegepant)(一种鼻内给药的gepant)的3期随机临床试验在主要结局指标(2小时无疼痛和无非最困扰症状)上证明了其疗效,且耐受性良好。这些gepants均未显示肝毒性证据。
CGRP是一种有效的血管扩张剂,并已知具有心脏保护作用,包括调节冠状血管炎症和血管舒张,通过影响心肌细胞减少心脏肥大,以及改善心肌收缩力和动脉血管舒张以预防系统性高血压。因此,抑制CGRP在理论上可能影响心脏健康;然而,值得注意的是,CGRP在心脏健康中的上述作用似乎是对损伤或损伤威胁的反应。在没有心脏损伤的情况下,CGRP不太可能对心脏健康产生不利影响,这反映在试验数据中,即使在乌布吉泮两项3期试验数据的高风险亚组中,心血管事件的发生率也未超过安慰剂。重要的是,这些试验排除了过去6个月内有急性心肌梗死、缺血性卒中或未控制高血压病史的患者。
过度使用许多偏头痛急性疗法,如曲普坦类药物,可能与药物过度使用性头痛有关,可能是由于影响了血清素能和内源性大麻素通路。迄今为止,gepants尚未被证明会导致患者出现药物过度使用性头痛,这与瑞美吉泮和阿托吉泮一致,因为它们在临床试验中已被证明对预防性治疗有效。Gepants提供了另一种靶向CGRP的方法,与CGRP mAbs并列。
PACAP最初在从羊下丘脑提取后以PACAP-38异构体形式被发现,并被发现能刺激cAMP。多年来,PACAP-38和PACAP-27的进一步作用已被阐明,包括抑制神经元凋亡、参与维持昼夜节律以及与情绪障碍(包括抑郁症)的关联。PAC1受体(PAC1-R)被归类为I型结合位点,对两种PACAP异构体的亲和力均显著高于VPAC1和VPAC2受体(VPAC1-R和VPAC2-R),后者对血管活性肠肽具有大致相等的亲和力。PAC1-R在结构上类似于降钙素家族受体,并且虽然在整个神经系统中广泛表达,但尤其集中在偏头痛相关区域,如下丘脑和脑桥核、脑干的迷走神经和上脊髓。发作间期的偏头痛患者已被证明其血浆PACAP-38免疫反应性低于健康对照受试者,而在发作期水平升高,这表明PACAP水平的波动程度可能在偏头痛生物学中起作用,而不仅仅是PACAP-38水平升高。PACAP-38在人血中半衰期短,主要与二肽基肽酶IV(DPP-IV)的降解有关;这个过程也被证明会释放出拮抗PAC1-R受体的代谢物。然而,使用DPP-IV抑制剂(而非激动剂)治疗与2型糖尿病患者慢性偏头痛发生率较低相关。这可能表明,PAC1-R受体激活的失调而非单纯该受体激活的增加可能在偏头痛病理生理学中起作用。
PACAP的神经保护作用被认为与增加的神经元兴奋性有关。在三叉神经痛的动物模型中,发现PACAP通过丝裂原活化蛋白激酶(MAPK)和细胞外信号调节激酶(ERK)通路调节NaV1.7。研究表明,输注PACAP-38可在与CGRP输注相似比例的偏头痛患者中诱发偏头痛头痛,但有趣的是,它还能在显著更高比例的患者中诱发前驱症状,这可能是因为下丘脑中PACAP-38的受体位点数量多于三叉神经节。一项关于AMG-301(一种PAC1-R抗体)在偏头痛患者中的临床试验未显示任何与安慰剂相比的显著治疗效果,但鉴于PAC1-R至少有12种剪接变体,用于靶向PAC1-R受体的表位可能不适用于相关变体。一种针对PACAP的mAb,Lu-AG09222,证明了与安慰剂相比每月平均偏头痛天数减少两天的统计学显著差异。对这些结果的一个可能解释是PACAP可能通过其对VPAC受体的作用而非仅通过PAC1-R诱发偏头痛;然而,值得注意的是,AMG-301试验招募的患者总体头痛天数较少。尽管上述剪接变异可能解释这一点,但另一种解释是PACAP可能通过另一个受体发挥诱发偏头痛的作用。例如,PACAP27和PACAP38都与G蛋白偶联受体55(GPR55)结合,导致GPR55内化。事实上,PACAP27是GPR55的任何内源性配体中效力最高的。GPR55是一种内源性大麻素样受体,遍布中枢神经系统,包括海马体、额叶、下丘脑、小脑和脑干结构(如导水管周围灰质),以及外周神经系统的脊髓神经元背根神经节。GRP55可能调节与偏头痛相关的多种功能,包括疼痛、情绪和代谢调节;然而,迄今为止,这基于动物模型中的证据。
PACAP激活肥大细胞受体MRGPRX2,该受体位于脑膜肥大细胞上,表明了另一种可能的作用机制。肥大细胞脱颗粒已被发现可激活脑膜伤害性感受器,可能促成偏头痛疼痛和行为,以及增强额外PACAP和CGRP的释放,鉴于PACAP引起的神经元兴奋性主要由PAC-1R受体内化后激活的MAPK/ERK通路驱动。事实上,对豚鼠受体的动物研究表明,通过抑制支架蛋白β-抑制蛋白或网格蛋白通路,可以阻止PAC-1R受体内化或内体形成,从而阻止PACAP对神经元兴奋性的调节。进一步的研究可能会更深入地了解PACAP肽或受体拮抗作用可能提供治疗益处的机制。
虽然CGRP受体由CLR和RAMP1蛋白组成,但CLR分别与RAMP2和RAMP3形成肾上腺髓质素1(AM1)和肾上腺髓质素2(AM2)受体。肾上腺髓质素也具有血管舒张特性,如CGRP,并定位于大鼠背根神经节神经元中,因此可能在疼痛感知中起作用。一项研究表明,在输注肾上腺髓质素后中位数4小时,可在无先兆偏头痛患者中诱发偏头痛发作。肾上腺髓质素也已被证明在CGRP受体上具有某些活性,尽管其在这些受体上的结合亲和力比CGRP低几个数量级。除了肾上腺髓质素,AM1受体也可能是一个靶点。RAMP2是AM1的蛋白质组分,与CGRP受体的蛋白质组分RAMP1相比,其在脑膜血管系统中的表达优先于冠状血管系统。因此,靶向AM1可能比靶向CGRP受体产生更少潜在心血管风险。AM2是CLR和RAMP3的异源二聚体,也被肾上腺髓质素激活,并且在动物模型中也由CGRP激活,尽管它可能具有不同的功能,特别是在骨骼健康和免疫系统中,并且可能避免其他靶点所具有的血管问题,实验研究中RAMP3敲除小鼠的存活与RAMP1和RAMP2敲除小鼠在子宫内死亡形成对比。鉴于CGRP与AM1和AM2受体之间的亲和力降低,以及肾上腺髓质素与CGRP受体之间的亲和力降低,目前尚不清楚靶向肾上腺髓质素的疗法是否必然同样有效,但它们可能作为现有CGRP拮抗剂的辅助或耐受性更好的替代品。
当降钙素受体(CTR)与RAMP1、RAMP2或RAMP3形成异源二聚体时,分别形成胰淀素1(AMY1)、胰淀素2(AMY2)或胰淀素3(AMY3)受体。AMY1已被证明与CGRP受体共定位于人三叉神经节中。普兰林肽(pramlintide)是一种已知的所有三种受体的激动剂,被证明可在88%的偏头痛患者中诱发偏头痛发作,而同一研究中97%的患者可由CGRP诱发偏头痛发作。同一研究表明,普兰林肽在CGRP和肾上腺髓质素受体上是弱激动剂,因此它可能通过胰淀素受体或肽拮抗以外的途径发挥这种作用。该研究未探讨胰淀素受体拮抗作用与CGRP受体拮抗作用结合可能产生的额外作用。胰淀素在胰岛素调节中也高度相关,胰淀素减少与糖尿病患者对葡萄糖摄入的胰岛素反应受损有关。这可能意味着胰淀素肽或受体拮抗作用存在胰岛素反应受损和饱腹感通路中断的风险。
尽管曲普坦类药物对偏头痛有效,但5HT1B受体的激动作用会增加冠状血管收缩的风险,从而产生心脏副作用。因此,曲普坦类药物由于其血管收缩特性而构成显著的脑血管和心血管风险,因此禁忌用于有血管风险因素的患者。选择性激动5HT1F受体(在整个脑实质中表达,包括在偏头痛相关结构中,但不在血管平滑肌中)将提供治疗益处并绕过血管问题。5HT1F受体也与腺苷酸环化酶负偶联,因此,激动该受体可以抑制中枢神经系统细胞的cAMP依赖性通路。正是基于这个原理,拉米地坦(lasmiditan)作为ditan类药物中的第一个药物被开发出来。拉米地坦在临床试验中已被证明作为偏头痛急性治疗有效,副作用包括头晕、嗜睡和恶心。一项荟萃分析表明,总体不良事件数量高于曲普坦类药物。
除了5HT1F受体,还有几个其他血清素通路靶点可能提供未来的治疗途径。丹麦的一项全基因组关联研究(GWAS)提示ADARB2中的一个位点与完全头痛自由相关。ADARB2仅存在于中枢神经系统,并在RNA编辑中起作用,包括编辑5HT2c受体。二氢麦角胺和阿戈美拉汀都曾用于治疗偏头痛,它们与5HT2c结合;然而,目前尚不存在专门针对该受体治疗偏头痛的疗法。一份报告指出,激活5HT2B受体可以抑制5HT1B受体功能,而5HT2B的这种功能本身可以被5HT2A受体的激活所抑制。这将5HT2受体的作用直接与一个已确立的偏头痛治疗通路联系起来。一项小型探索性双盲安慰剂对照试验报告,单剂量裸盖菇素(激活5HT2A受体)在随后2周内减少了偏头痛天数。然而,缺乏更大规模的研究来支持这些结果,并且裸盖菇素也靶向一系列其他血清素受体,导致众所周知的致幻副作用。
5HT1A受体在血清素能张力调节中起关键作用;中缝核中这些受体的突触前激活导致超极化,从而抑制血清素能神经元。这些受体在非血清素能神经元中也作为异源受体发挥作用,并可能调节中枢神经系统其他区域的其他神经递质释放,包括蓝斑中去甲肾上腺素的释放。这进一步得到了研究的证实,研究表明丁螺环酮(一种5HT1A激动剂)可在人类中引起剂量依赖性瞳孔缩小。
5HT1A受体对不同区域中枢神经系统中神经递质释放的调节作用,特别是脑干,可能有助于偏头痛病理生理学。一项正电子发射断层扫描研究显示,与健康对照受试者和未触发发作的偏头痛患者相比,气味触发偏头痛发作的患者在脑桥中缝核中5HT1A拮抗剂放射性示踪剂的结合增加,表明5HT1A受体的异常活动可能发生在偏头痛头痛期的早期。另一项研究进一步阐述了这种潜在作用,表明无先兆偏头痛患者在服用丁螺环酮后催乳素升高幅度高于年龄和月经周期匹配的健康对照受试者,突出了偏头痛患者5HT1A受体相对超敏感性的可能性。几种现有的偏头痛预防性药物,如阿米替林、普萘洛尔和丙戊酸钠,影响5HT1A受体;然而,目前尚无单独特异性靶向该受体的疗法,并且上述药物的偏头痛预防作用机制可能通过其他通路。
TRPA1和TRPV1是属于更广泛的瞬时受体电位(TRP)家族的离子通道,调节外周神经系统对机械和化学应激物的疼痛反应,尽管它们在中枢神经系统中也被发现,但其作用仍不清楚。TRP通道的激活已被证明会导致CGRP的释放。已知的偏头痛诱发因素,包括酒精,会激活TRP通道。然而,一项TRPV1拮抗作用的2期临床试验证明其疗效劣于安慰剂。多项GWAS研究已识别出TRPM8基因上的位点可能对偏头痛有保护作用,该基因编码另一个离子通道。TRPM8也与睾酮相互作用,在偏头痛小鼠模型中,雄性小鼠比雌性小鼠从超敏反应中恢复得更快,这可能是由于雄性小鼠中TRPM8的表达或活性增加,这可能为女性偏头痛患病率高于男性提供了一种解释。局部应用薄荷醇(一种有效的TRPM8激活剂)已被证明可以治疗偏头痛;然而,鉴于TRPM8在中枢神经系统中的作用尚未被表征,需要进一步研究以确定通过替代给药途径进行更靶向的治疗是否可能更有益。其他TRP通道,如TRPM3,也在三叉神经节中表达,并在动物模型中与伤害性信号传导有关;然而,TRPM3拮抗剂BHV-2100最近在一项Biohaven投资者会议上报告了失败的2a期研究。
由神经肽与CGRP、肾上腺髓质素和胰淀素受体结合引发的细胞内cAMP信号级联导致ATP敏感性钾通道(称为KATP)的开放。KATP通道的关键作用通过研究得到强调,研究表明输注左克罗卡林(levcromakalim)(一种有效的KATP通道开放剂)可在偏头痛个体中引发偏头痛样头痛。然而,值得注意的是,另一项研究中健康对照受试者在左克罗卡林后头痛发生率也很高,这表明KATP通道可能在一般性头痛中起作用,而非作为偏头痛的特异性通路。鉴于KATP通道分布广泛,使用拮抗剂时存在其他脱靶效应的潜力,例如阻断存在于大脑和胰腺中的Kir6.2/SUR1亚型会导致低血糖,以及阻断存在于血管平滑细胞中的Kir6.1/SUR2亚型会导致高血压。没有证据表明输注Kir6.2/SUR1亚型开放剂会诱发偏头痛,这表明参与偏头痛病理生理学的KATP通道亚型是血管性的。然而,给予格列本脲(glibenclamide)(一种两种亚型的拮抗剂)并未抑制由左克罗卡林或CGRP引起的头痛,尽管这可能是由于较高剂量下Kir6.2/SUR1的低血糖不良反应限制了有效阻断Kir6.1/SUR2的剂量。因此,更特异性地阻断Kir6.1/SUR2可能是未来研究的一个潜在治疗途径,但也携带显著潜在风险,因为该通道亚型在冠状血管的血管平滑肌细胞中高表达。
硝酸甘油(GTN)[硝化甘油(NTG)]长期以来已知会引发头痛。GTN输注已广泛用于偏头痛的实验模型,并已被证明可可靠地触发偏头痛患者的偏头痛头痛和前驱症状。GTN是一种一氧化氮(NO)供体和有效的血管扩张剂,在5分钟内达到最大血管舒张效果;然而,这些研究表明偏头痛头痛在给药后数小时才发作,这表明血管舒张本身不足以解释这种诱发模型。事实上,NO在氧化应激中起关键作用,但也激活sGC-cGMP信号转导,后者在中枢神经系统中充当第二信使系统。三种一氧化氮合酶(NOS)亚型内源性催化NO:内皮型NOS(eNOS)、诱导型NOS(iNOS)和神经元型NOS(nNOS)。
一项关于L-NMMA(一种非选择性NOS抑制剂)的安慰剂对照试验在治疗急性偏头痛发作方面有效。然而,据报道有显著的并发高血压和心动过缓的不良反应,可能是通过抑制eNOS亚型实现的,该亚型在血管内皮中大量表达。一项关于GW274150(一种选择性iNOS抑制剂)的2期临床试验未能证明其在偏头痛预防上优于安慰剂,在急性发作研究中也无效。这可能归因于iNOS主要在免疫调节和血管张力中起作用,在神经系统结构中表达极少。
nNOS亚型因其在偏头痛相关结构中的表达而引起了极大兴趣,这些结构包括外周神经系统(如三叉神经节)和中枢神经系统(如下丘脑、海马体、丘脑和脑桥),它在调节随时间变化的突触传递中起作用,包括通过调节GABA能和代谢型谷氨酸受体。非特异性NOS抑制剂N(G)-硝基-L-精氨酸甲酯(L-NAME)已被证明可减少三叉颈神经元中Fos蛋白的表达。尽管nNOS特异性抑制剂S-甲基-L-硫代瓜氨酸(SMTC)可以减少神经源性硬脑膜血管舒张,但nNOS可能是一个更可行的治疗靶点,因为其在血管内皮中的表达相对于eNOS较少。虽然本身不在心脏组织中表达,但nNOS已被证明对血管张力有影响,使用SMTC抑制会导致前臂基础血流量减少以及脑血流量减少。此外,在动物模型的下丘脑中阻断nNOS会导致系统性高血压,可能是由于扰乱了自主神经功能。目前,尚无研究专门探讨nNOS抑制在偏头痛患者中的作用,尽管一项关于联合nNOS阻断剂/曲普坦药物NXN-188的研究在急性偏头痛和一项偏头痛伴先兆的试验中失败了。虽然第一项研究未显示2小时无疼痛的统计学显著反应,但它确实证明了在4至24小时之间有反应,并且在1小时时与安慰剂相比完全缓解了所有偏头痛症状。后一项研究有一个显著的局限性,即由于大多数患者发作频率太低而无法纳入分析,导致样本量不足以得出结论。
突触后密度蛋白95(PSD-95)是一种支架蛋白,与nNOS偶联后,与NMDA受体结合。值得注意的是,PSD-95可以通过与nNOS偶联以外的机制与NMDA受体结合;例如,它也可能与低密度脂蛋白受体相关蛋白1(LRP1)偶联以结合NMDA受体,尽管这迄今为止仅在动物模型而非人体组织中得到证明。然而,这可能特别令人感兴趣,因为GWAS研究已显示LRP1基因与偏头痛之间存在关联。PSD-95在慢性疼痛模型中被涉及,一项研究证明,在慢性偏头痛小鼠模型的三叉神经核中,PSD-95的过度表达与催产素受体共定位。在同一项研究中,鼻内催产素减弱了PSD-95的过度表达,抑制了下游cAMP-PKA-CREB信号通路,并减轻了NTG诱导的痛觉过敏。催产素在一些试验中显示出作为偏头痛治疗的前景,尽管一项更大的试验未能达到其主要终点,部分原因是研究队列中的安慰剂反应高于预期。Nerinetide是一种二十肽,与PSD-95结合,从而干扰其功能,并在一项用于卒中的试验中被证明对人类是安全的;它迄今尚未进行偏头痛试验。与通过NOS靶向不同,nerinetide治疗未伴随血压升高。PSD-95由于其更选择性地在神经组织中表达,提供了nNOS之外的替代选择,尽管临床数据仍处于初期阶段。
偏头痛治疗领域持续发展,CGRP mAbs在过去十年中增强了治疗这种情况的手段,为许多慢性头痛患者提供了益处,拉米地坦也被引入作为急性治疗选择,特别适用于可能对其他药物有禁忌症的患者。然而,许多患者的偏头痛发作对这些疗法无反应,因此仍需开发靶向替代通路的新疗法。一种PACAP mAb在早期2期临床试验中已被证明有效,并可能作为偏头痛治疗的另一种选择,其他潜在靶点如胰淀素、肾上腺髓质素、离子通道和nNOS正在进行临床前研究。偏头痛临床表现的异质性以及个体间对偏头痛预防性药物反应的可变性可能表明潜在疾病生物学的差异,也许偏头痛治疗的未来将侧重于精准医疗,为患者提供针对其特定疾病生物学中每种通路的定制治疗,而非单一的通用治疗途径。
生物通微信公众号

