
-

生物通官微
陪你抓住生命科技
跳动的脉搏
生物通官微
陪你抓住生命科技
跳动的脉搏
生命学院陈浩东课题组综述植物如何感受重力
【字体: 大 中 小 】 时间:2026年06月10日 来源:清华园生命学院
编辑推荐:
重力是地球上最恒定的物理环境因子之一。对动物而言,重力主要影响身体在空间中的姿态控制、平衡维持和运动方式;而对无法移动的植物来说,重力则从根本上塑造了其生长方向和形态结构。一般而言,植物的茎向上生长,根向下生长,侧枝和侧根则以特定角度展开,共同构成植物适应环境、获取光照和水分养分的空间架构。一个有趣的问题是:植物究竟如何“知道”重力来自哪个方向?更进一步,植物细胞如何将这一非接触性的物理力,转...
重力是地球上最恒定的物理环境因子之一。对动物而言,重力主要影响身体在空间中的姿态控制、平衡维持和运动方式;而对无法移动的植物来说,重力则从根本上塑造了其生长方向和形态结构。一般而言,植物的茎向上生长,根向下生长,侧枝和侧根则以特定角度展开,共同构成植物适应环境、获取光照和水分养分的空间架构。一个有趣的问题是:植物究竟如何“知道”重力来自哪个方向?更进一步,植物细胞如何将这一非接触性的物理力,转化为细胞内可识别、可传递、并最终调控生长方向的生理信号?
2026年6月4日,清华大学陈浩东课题组应邀在《新植物学家》(New Phytologist)上发表 Tansley Review,题为“植物的重力感受”(Gravity sensing in plants)。该综述系统总结了植物重力感受研究的经典理论、最新分子机制、演化线索以及农业应用前景,并特别强调了近年来该领域的关键突破:淀粉体沉降能够重塑LAZY蛋白的细胞极性,从而为已有百余年历史的“淀粉-平衡石”假说提供了明确的分子解释。
1.重力感受细胞:植物在哪里感受重力?
植物向重力性的过程可划分为三个连续步骤:重力感受(gravity sensing/perception)、信号转导(signal transduction)和生长反应(growth response)。重力感受又可以分为物理感受(susception)与生理信号转换(physiological signal conversion)两个阶段。重力作用于植物的每一个细胞,但并不是所有细胞都能感受重力。真正承担重力感受功能的是一类特化细胞,称为重力感受细胞(statocytes)。
在根中,主要的重力感受细胞位于根冠的柱细胞(columella cells)。早在达尔文父子的经典著作The Power of Movement in Plants中,他们就提出根尖能够感受外界信号,并将某种“影响”传递给相邻区域,引导根发生弯曲。后续去除根冠、激光消融根冠细胞以及遗传学研究进一步证明,根冠柱细胞是根向重力性反应的核心感受部位。
在地上部分,茎和下胚轴的重力感受主要发生在内皮层细胞(endodermal cells)中。拟南芥中缺失正常内皮层形成的突变体会丧失茎的向重力性,而根的向重力性不受影响,说明根和茎使用不同的细胞类型感受重力。文章还进一步讨论了其他潜在的重力感受细胞,例如玉米去根冠后仍可保留一定向重力反应,石松类和蕨类植物根中的淀粉体分布也与种子植物明显不同,提示植物界可能存在多样化的重力感受方式(图1)。
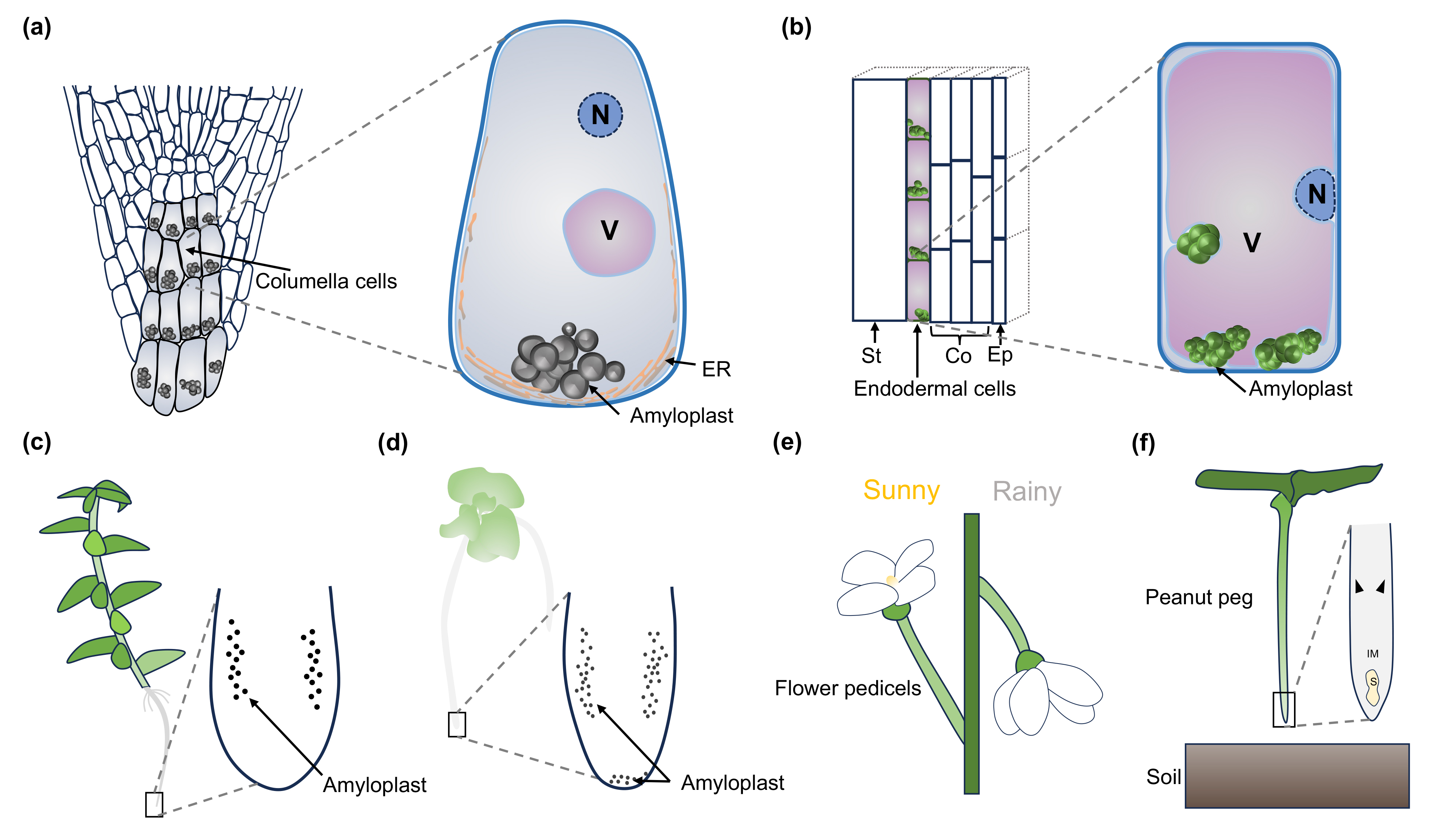
图1. 维管植物中的重力感受细胞
2. 亚细胞结构:谁参与了重力感受?
在重力感受细胞内部,参与重力感受的最经典、最核心细胞器是淀粉体(amyloplast)。淀粉体富含淀粉颗粒,密度高于周围细胞质,因此能够沿重力方向快速沉降。这一现象构成了“淀粉-平衡石”假说的物理基础。
不过,淀粉体并不是孤立发挥作用。综述指出,细胞骨架、液泡、质膜和内质网等亚细胞结构都可能参与重力感受过程。微丝骨架可以限制或调控淀粉体沉降,因此不同药物处理、不同发育阶段或不同器官中,细胞骨架对向重力性的影响可能出现差异。液泡在茎内皮层细胞中特别重要,因为大型中央液泡会影响淀粉体运动,而根冠柱细胞中液泡较小,其影响相对有限。
质膜则是近年来研究的关键焦点。淀粉体沉降后,LAZY蛋白会重新定位到细胞新的下侧质膜区域,从而把淀粉体的物理位置信息转化为细胞极性信息。与此同时,质膜上的机械敏感离子通道、钙信号以及内质网-质膜接触位点也被认为可能参与重力信号的感受和转导,但这些机制仍有待进一步验证。
3. 分子机制:淀粉体“带着”LAZY建立细胞极性
本综述最核心的部分,是对植物重力感受分子机制的系统总结。过去一个多世纪中,科学界普遍接受淀粉体沉降参与重力感受,但淀粉体如何真正触发信号,一直缺乏清晰的分子解释。
近年来,综述作者团队及国际同行的突破性研究表明,LAZY蛋白是连接淀粉体沉降与生长反应的核心分子。LAZY基因最早因玉米和水稻中“趴伏生长”的突变体而得名。之后,LAZY家族成员被证明广泛参与植物茎的负向重力性、根的正向重力性以及侧枝、侧根角度调控。
在拟南芥根冠柱细胞中,LAZY2、LAZY3和LAZY4在重力刺激后会重新分布到细胞新的下侧质膜。更重要的是,这一过程依赖淀粉体沉降:淀粉体带动LAZY蛋白靠近新的下侧质膜,LAZY蛋白随后在该区域形成极性富集。换句话说,细胞通过“淀粉体沉降―LAZY极性重塑”这一过程,将重力方向这一物理信息转化为细胞内的生理信号。
文章进一步总结了这一过程的分子细节:MKK5-MPK3激酶模块可促进LAZY蛋白磷酸化;磷酸化后的LAZY蛋白通过与TOC复合体成员相互作用定位到淀粉体表面;当淀粉体沿重力方向沉降时,LAZY蛋白被带到新的下侧质膜,并可能通过碱性-疏水区域与膜磷脂结合。由此,百年前提出的“淀粉-平衡石”假说终于获得了分子层面的清晰解释(图2)。
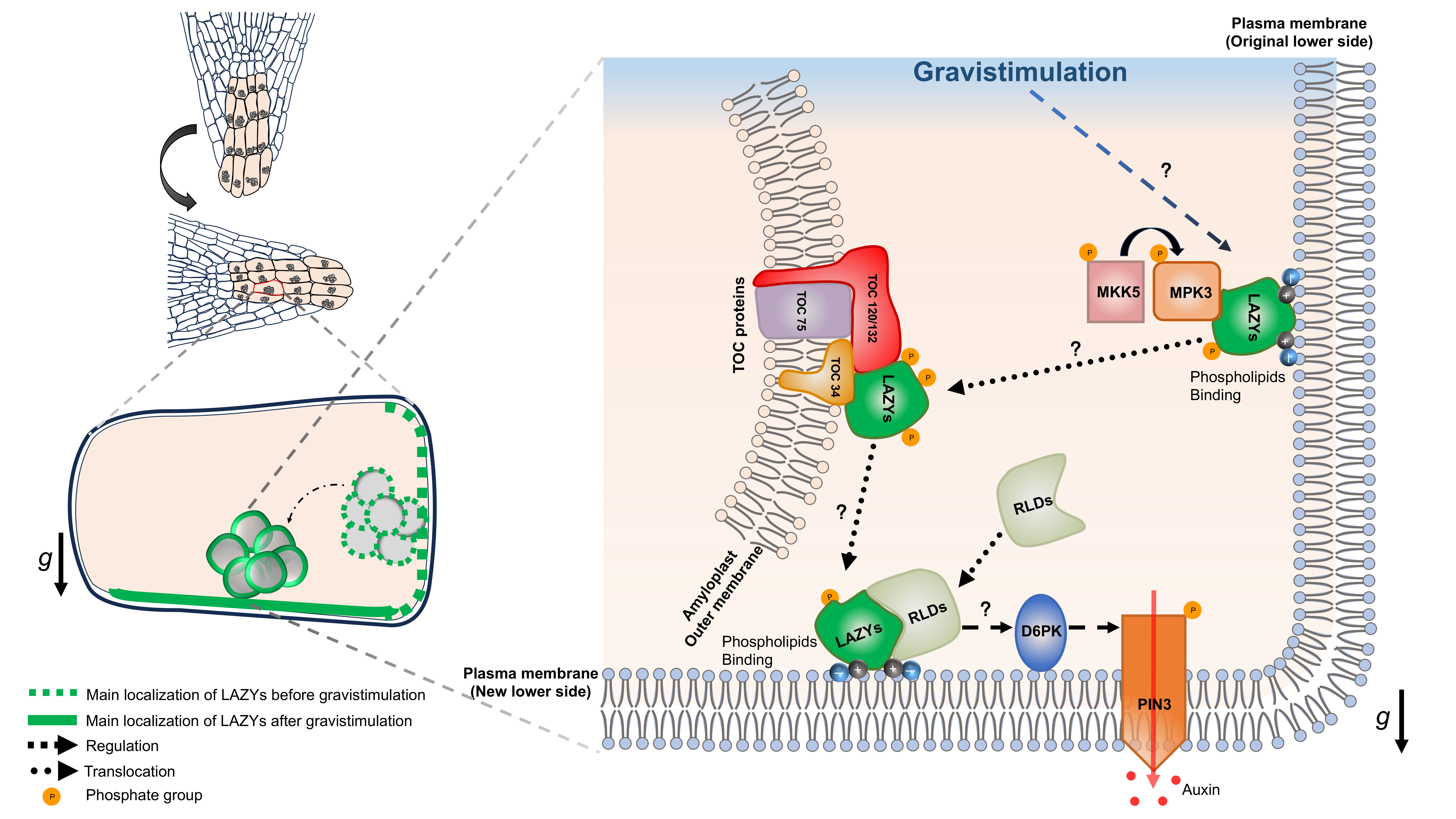
图2. 淀粉体沉降重塑 LAZY 蛋白极性以实现重力感受
4. 信号转导:从LAZY极性到生长弯曲
重力感受完成后,植物还需要把局部细胞极性转化为整个器官的不对称生长。这里的核心信号是生长素(auxin)。经典的Cholodny-Went学说认为,植物向性弯曲源于生长素的不对称分布。综述指出,在根和茎中,重力刺激后生长素都会更多积累在器官下侧,但两者产生的生长反应却相反:茎向上弯曲,根向下弯曲。这是因为不同器官对生长素的响应方式不同,但具体机制仍有待进一步研究。
在重力感受细胞内,LAZY蛋白极性分布后可招募RLD、D6PK等因子,并进一步影响PIN3、PIN7等生长素输出载体的极性定位,从而建立生长素不对称运输(图2)。在根中,AUX1和PIN2继续将生长素信号从根冠传递到伸长区,使下侧生长受到更强的抑制,最终导致根向下弯曲。在茎或下胚轴中,生长素则可通过ARF7/ARF19和SAUR等早期响应基因促进下侧细胞伸长,从而推动地上器官向上弯曲。
文章还特别强调,向重力性反应不仅需要启动,也需要终止。若植物持续弯曲而无法停止,就会出现过度弯曲。已有研究表明,生长素可反馈调节PIN3极性,使生长素分布重新对称化,从而参与弯曲终止。然而,生长素梯度消失之后,器官如何通过整合细胞壁力学与微管重排等过程继续调整形态,并精准控制弯曲的终止位置,仍是未来需要解决的重要问题。
5. 非维管植物:从演化角度理解重力感受
植物重力感受并非种子植物独有。苔藓、轮藻等非维管植物同样表现出向重力性,这为理解重力感受的演化提供了独特窗口。
在小立碗藓和角齿藓等藓类植物中,拟茎体可以表现负向重力性,假根表现正向重力性;在黑暗条件下,苔藓原丝体中的轴丝体也可表现明显的负向重力性。与维管植物不同,苔藓原丝体常在单个顶端生长细胞中同时完成重力感受和生长响应,因此其机制可能更依赖细胞骨架和顶端生长调控(图3)。
地钱也具有明显的向重力性。在黑暗条件下,地钱叶状体可形成向上生长的狭长结构,其中的储藏组织细胞可观察到淀粉体沉降(图3)。淀粉缺失突变体的向重力性减弱但并未完全消失,说明淀粉体对地钱完整的重力反应十分重要。
轮藻的假根和原丝体则提供了另一个极具启发性的模型。它们是单细胞系统,能够通过平衡石与特定质膜区域接触来感受重力,并通过钙信号、肌动蛋白、胞吐和顶体等结构调控生长方向。比较这些不同植物类群,有助于回答一个更深层次的问题:植物在登陆和演化过程中,如何逐步建立从单细胞到多细胞、从局部感受扩展到器官弯曲的重力响应系统?
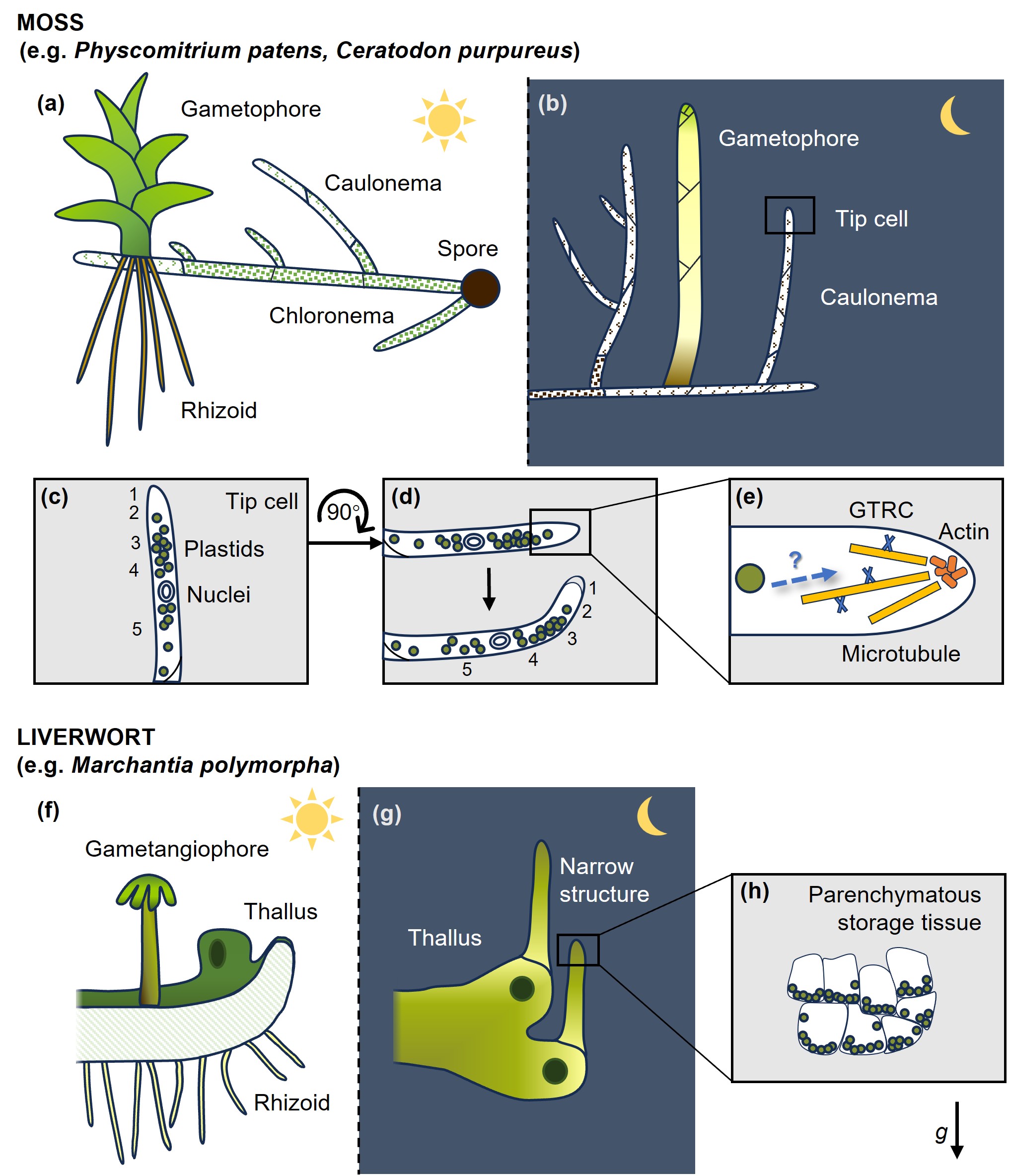
图3. 苔藓植物中的向重力性
6. 未知重力感受机制:植物解读物理信息的新方式
尽管“淀粉体沉降―LAZY极性重塑”已经成为当前解释种子植物重力感受的重要模型,但综述也指出,植物中可能存在独立于经典淀粉体机制的替代重力感受途径。例如,去除根冠的玉米根仍可缓慢弯曲;石松和蕨类植物根中淀粉体并不表现典型沉降。这些现象提示,植物可能拥有多重、重叠的重力感受系统。与此同时,LAZY是否是所有重力信号的必要组成部分,还是主要作为经典淀粉体途径中的重力信号正向转导因子,仍有待澄清。近期关于SLQ1突变可抑制lazy高阶突变体表型的研究,也暗示了LAZY非依赖性重力信号通路的存在。
解析这些尚未被定义的重力感受机制,不仅是对经典淀粉体模型的补充,更可能揭示植物如何将非接触性的物理力转化为生理信号的普适原理。未来,结合正向遗传筛选、跨类群比较和多组学分析,有望发现新的重力感受因子,并进一步重构植物重力感受的完整网络。
7. 农业应用:从重力感受机制到作物架构优化
在农业应用方面,重力感受机制直接关系到作物理想株型设计。地下部分,水稻DRO1可增强根向重力性,使根系扎得更深,从而提高干旱条件下对深层水分的获取能力;相反,在盐碱或低磷环境中,较浅的根系结构有时更有利于作物适应表层土壤资源(图4)。地上部分,LAZY1、TAC1等基因调控分蘖角、枝条角度和树冠结构,影响适宜密植程度、机械化采收效率和产量等。近期大豆中TAC1同源基因的研究也表明,改变侧枝角度可提高植株紧凑性,提高密植条件下的产量。
这些研究表明,重力感受及其下游信号通路中的关键基因,是改良作物根系结构、株型紧凑性和环境适应性的重要靶点。随着机制解析与精准基因编辑技术的发展,植物向重力性研究有望为培育高产、抗逆并适应未来气候变化的理想作物提供新的理论基础和分子工具。
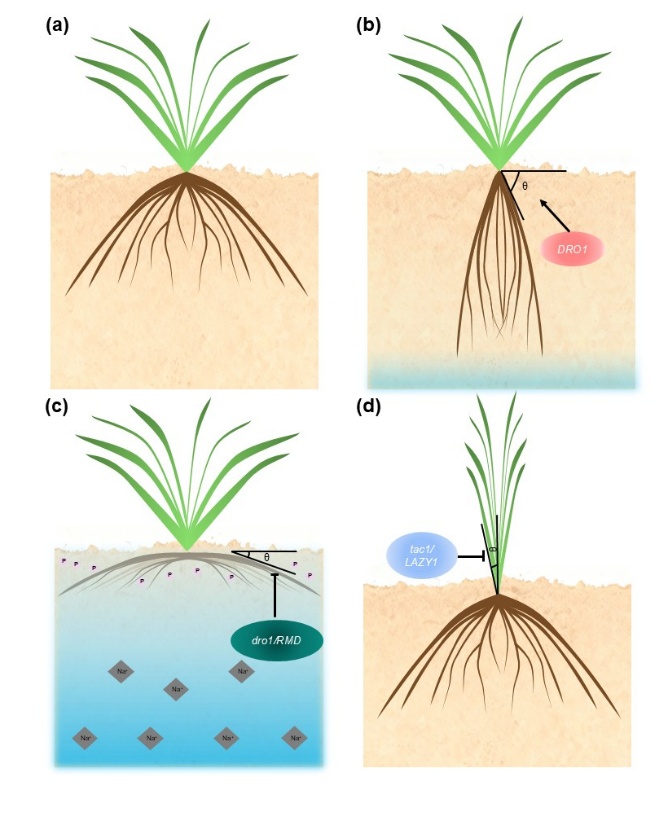
图4. 重力感受相关基因对水稻株型的影响
8. 未来展望
在综述最后,作者提出了植物重力感受领域未来需要重点解决的几个关键问题。首先,种子植物中LAZY蛋白如何在质膜和淀粉体外膜之间精确转运,仍缺乏完整机制解释。其次,重力刺激后LAZY蛋白的磷酸化由哪些上游事件触发、上侧质膜上的LAZY是否被降解、LAZY如何进一步调控PIN介导的生长素运输,都是需要深入研究的核心问题。第三,除经典淀粉体途径外,植物是否存在其他的重力感受系统,将是未来可能带来概念突破的重要方向。
作者强调,植物向重力性研究不仅是理解植物形态建成的基础科学问题,也为作物理想株型、根系结构优化、抗逆育种和空间农业提供了理论基础。未来,通过遗传学、多组学、细胞生物学、生物物理学以及人工智能等交叉手段,研究者有望识别更多重力感受与信号转导因子,并将这些知识用于精准设计适应不同环境的作物构型。
这篇综述由清华大学生命科学学院、绿色生物制造全国重点实验室、清华-北大生命科学联合中心陈浩东课题组完成。陈浩东副教授为论文通讯作者,王笑连博士(曾为陈浩东课题组博士后,现为英国布里斯托大学博士后)为论文第一作者,2022级博士研究生蔡心悦、博士后霍寰为论文共同作者。该工作得到了国家自然科学基金和清华大学自主科研计划支持。
原文链接:https://doi.org/10.1111/nph.71324

